Imprimer
Imprimer
ph. A.S. Walker

Examen d'une culture de spores comme celle de la photo en médaillon. La technique est plus lourde que la PCR mais détecte toutes les résistances même celles dues à des mécanismes encore inconnus.
La résistance aux fongicides est, on le sait, un phénomène courant chez les champignons phytopathogènes c'est-à-dire agents de maladies des plantes. C'est le cas en particulier de Septoria tritici, agent de la septoriose, principale maladie du blé en France. De nombreux isolats de S. tritici résistant à divers fongicides sont détectés et généralisés depuis de nombreuses années. Plus récemment, différents partenaires ont détecté de nouveaux isolats encore plus résistants aux fongicides IDM... Mais les programmes fongicides incluant ces substances restent efficaces dans la grande majorité des cas. Comment expliquer cette apparente contradiction ? Quels enseignements tirer de 2010 ? Faut-il prendre en compte l'évolution récente des résistances dans les stratégies de traitement ? Réponses ci-après.
Photo en médaillon : vue au microscope de spores de S. tritici prélevées en 2010, après incubation en présence de fongicide pharmaceutique à haute dose. La population contient 50 % de phénotypes MDR, dont les filaments croissent autant ici que dans le témoin sans fongicide.
La pression parasitaire sur céréales a été faible en 2010, en particulier celle de la septoriose, mais la campagne n'en fut pas moins riche d'enseignements. Certes, il fut parfois difficile de mener à terme les essais au champ, en particulier dans la moitié nord de la France où le déficit hydrique fut plus intense, mais de nombreux échantillons de blé « septorié » ont pu malgré tout être fournis à l'INRA Bioger pour en analyser les résistances aux fongicides.
La résistance biologique d'un isolat fongique se caractérise par sa capacité à être moins sensible à une dose de fongicide qui, normalement, devrait le contrôler. Ce caractère est transmissible à sa descendance puisqu'il est déterminé génétiquement. Cela implique qu'une ou plusieurs modifications dans le génome du champignon ont été sélectionnées et lui ont permis d'acquérir ce nouveau caractère. Au niveau des populations, on parlera de résistance en pratique lorsque l'on observe une baisse de l'efficacité des traitements, liée à la présence de souches résistantes dans les populations.
Méthodologie
Depuis une dizaine d'années, l'INRA Bioger analyse des populations de septoriose issues d'essais menés par la profession. Les échantillons de feuilles de blé malades sont utilisés pour constituer des suspensions de spores en mélange. Celles-ci sont inoculées in vitro sur des milieux nutritifs contenant des doses discriminantes de fongicides, ie des doses pour lesquelles seuls les individus résistants à la molécule utilisée peuvent croître.
Ce test permet in fine d'estimer la proportion de chaque type de résistance dans l'échantillon, et ce à partir d'un grand nombre de spores, ce qui constitue un échantillon représentatif de la parcelle.
En 2010, plus de 700 populations de S. tritici ont ainsi été analysées.
Certaines souches ont été isolées (monospores) pour caractériser plus finement leur phénotype (CI50, spectres de résistance croisée).
Trois situations stables
Vis-à-vis des benzimidazoles
En 2010, la fréquence moyenne des souches résistant spécifiquement aux benzimidazoles (ex : carbendazime, thiophanate-méthyl) est de 87 %. Ainsi, cette résistance reste généralisée dans les populations depuis au moins une dizaine d'années alors que l'usage de cette famille sur septoriose est anecdotique.
C'est la preuve d'une « absence de coût » ou d'un coût réduit, liés à cette résistance affectant le cytosquelette (tubuline) des champignons. Autrement dit, ces souches n'ont pas « payé » l'acquisition de la résistance aux benzimidazoles par l'affaiblissement d'une fonction vitale (laquelle ferait diminuer leurs populations en l'absence de traitement, au bénéfice des souches sensibles).
Vis-à-vis des strobilurines
La résistance aux strobilurines (ex : azoxystrobine, picoxystrobine, trifloxystrobine…), détectée en France en 2002, n'a cessé d'augmenter en fréquence depuis cette date. Elle est généralisée depuis quelques années même dans le sud de la France. Sans surprise, 99 % des populations testées présentaient encore plus de 50 % d'isolats résistants aux strobilurines en 2010, malgré un usage qui se réduit sur septoriose.
À noter que le mécanisme de cette résistance (changement G143A dans la protéine du cytochrome b, intervenant dans la respiration cellulaire) est largement répandu chez plus de 37 champignons phytopathogènes dans le monde (Leroux and Walker, 2010b).
Vis-à-vis des SDHI
Les inhibiteurs de la succinate deshydrogénase (SDHI) sont constitués de plusieurs sous-familles, dont certaines, représentées par des fongicides comme la carboxine ou l'oxycarboxine, sont connues depuis les années 70.
Les SDHI de nouvelle génération appartiennent quasiment tous à la sous famille des pyridinecarboxamides (ex : boscalid, seul SDHI actuellement autorisé sur céréales en France) ou des N-méthylpyrazoles-carboxamides (ex : bixafen, isopyrazam, penthiopyrad, sedaxane… ; ces SDHI seront très prochainement introduits sur le marché) (Leroux et Walker, 2010a).
Bien que des résistances à ces fongicides soient détectées chez d'autres champignons phytopathogènes, aucune résistance spécifique aux SDHI n'a été caractérisée à ce jour chez des isolats du champ de S. tritici.
Sensibilité aux IDM
Rappel sur une situation complexe
Les inhibiteurs de la déméthylation des stérols (IDM) constituent la base de la protection fongicide contre la septoriose, en particulier depuis la généralisation de la résistance aux strobilurines. Les IDM sont représentés majoritairement par les triazoles (ex : cyproconazole, époxiconazole, propiconazole, tébuconazole…), mais également par un imidazole (prochloraze) et une triazolinethione (prothioconazole).
La résistance aux IDM est complexe puisque, à l'inverse des exemples précédents, plusieurs phénotypes de résistance ont été sélectionnés au cours du temps et sont présents en mélange dans les populations.
Ces isolats se distinguent d'une part par leur niveau de résistance (ie intensité de la résistance) aux fongicides, et d'autre part par leur spectre de résistance croisée (ie leur capacité à résister simultanément à plusieurs molécules).
Ainsi, jusqu'en 2007, notre laboratoire avait caractérisé jusqu'à 8 phénotypes différents (nommés TriR1 à TriR8), qu'il était possible de regrouper en 5 phénotypes globalement légèrement résistants aux IDM (TriR1 à TriR5 ou TriLR) et 3 phénotypes globalement moyennement résistants aux IDM (TriR6 à TriR8 ou TriMR) (Tableau 1) (Leroux et al., 2007)
Depuis le milieu des années 2000, ces isolats TriMR, en particulier les isolats TriR6, sont largement dominants dans les populations françaises (plus de 50 % en fréquence dans 96 % des populations échantillonnées ; Figure 1) mais aussi européennes (Stammler et al., 2008).
La résistance de ces isolats est principalement déterminée par des altérations du gène cyp51, qui code pour la protéine 14α-déméthylase, cible des IDM. En particulier, la mutation I381V, sélectionnée au début des années 2000, est identifiée chez tous les phénotypes TriMR (Figure 2 page suivante).
L'apport, en terme de résistance, des différentes mutations affectant cyp51 a récemment été quantifié par des études d'expression hétérologue chez la levure (Cools et al., 2010).
2008, détection des phénotypes émergents
Depuis 2008, de nouvelles souches, plus fortement résistantes aux IDM, sont détectées dans les populations françaises. Ces souches ne sont pas homogènes entre elles et correspondent à plusieurs phénotypes, dits phénotypes émergents (Tableau 1 ; Figure 2) (Leroux et Walker, 2011) :
– Les souches TriR5+ et TriR8+ possèdent un spectre de résistance proche de celui des souches TriR5 et TriR8 respectivement, mais sont plus résistantes. Elles présentent les mêmes mutations dans le gène cyp51 que les souches dont elles dérivent. Un mécanisme additionnel est probablement mis en œuvre chez ces isolats ; il reste à l'identifier.
– Les souches TriR9, TriR10 et TriR11 sont fortement résistantes à un ou quelques IDM. Elles sont globalement plus résistantes que les souches TriMR décrites jusqu'à présent. Leur spectre de résistance est nouveau. L'analyse du polymorphisme du gène cyp51 met en évidence des mutations déjà connues (ex : S524T, D134G) à faible fréquence et nouvellement combinées avec des mutations largement répandues. Ainsi, il est probable que le phénotype TriR9, largement répandu en Irlande, ait évolué à partir des isolats TriR5, fortement implantés dans ce pays. En 2010, en sus des souches décrites dans le tableau 1, une grande variété de phénotypes présentant de nouvelles combinaisons de mutations déjà connues a été détectée dans divers pays, conduisant à élargir la variété des génotypes de cyp51. Si le mécanisme de mutation de cible semble prépondérant chez ce type d'isolat, il est probable que la capacité d'évolution du gène cyp51 soit encore sous-estimée.
– Enfin, les souches MDR6, MDR7 et MDR10 présentent des niveaux de résistance très forts à tous les IDM et au prochloraze. Ces isolats présentent les mêmes mutations dans cyp51 respectivement que les souches TriR6, TriR7 et TriR10 dont les niveaux de résistance sont globalement moyens. Les souches MDR possèdent donc un mécanisme de résistance additionnel. Des travaux complémentaires ont montré qu'elles peuvent excréter plus efficacement les molécules toxiques, fongicides en particulier, hors de la cellule fongique, grâce à des pompes membranaires (de type ABC ou MFS transporteur) présentes en plus grande quantité dans ces isolats. Cette surexpression probable de transporteurs membranaires caractérise le mécanisme dit de MDR (MultiDrug Resistance), largement connu dans le domaine médical pour le cas de la résistance aux anticancéreux par exemple. La spécificité relative des transporteurs impliqués dans le phénotype MDR des souches de S. tritici explique probablement que ces souches présentent une résistance croisée positive avec les QoI (dont font partie les strobilurines), mais aussi avec les SDHI, avec cependant de faibles niveaux de résistance (de l'ordre de 5 à 15 selon les molécules et les phénotypes).
Évolution de 2008 à 2010
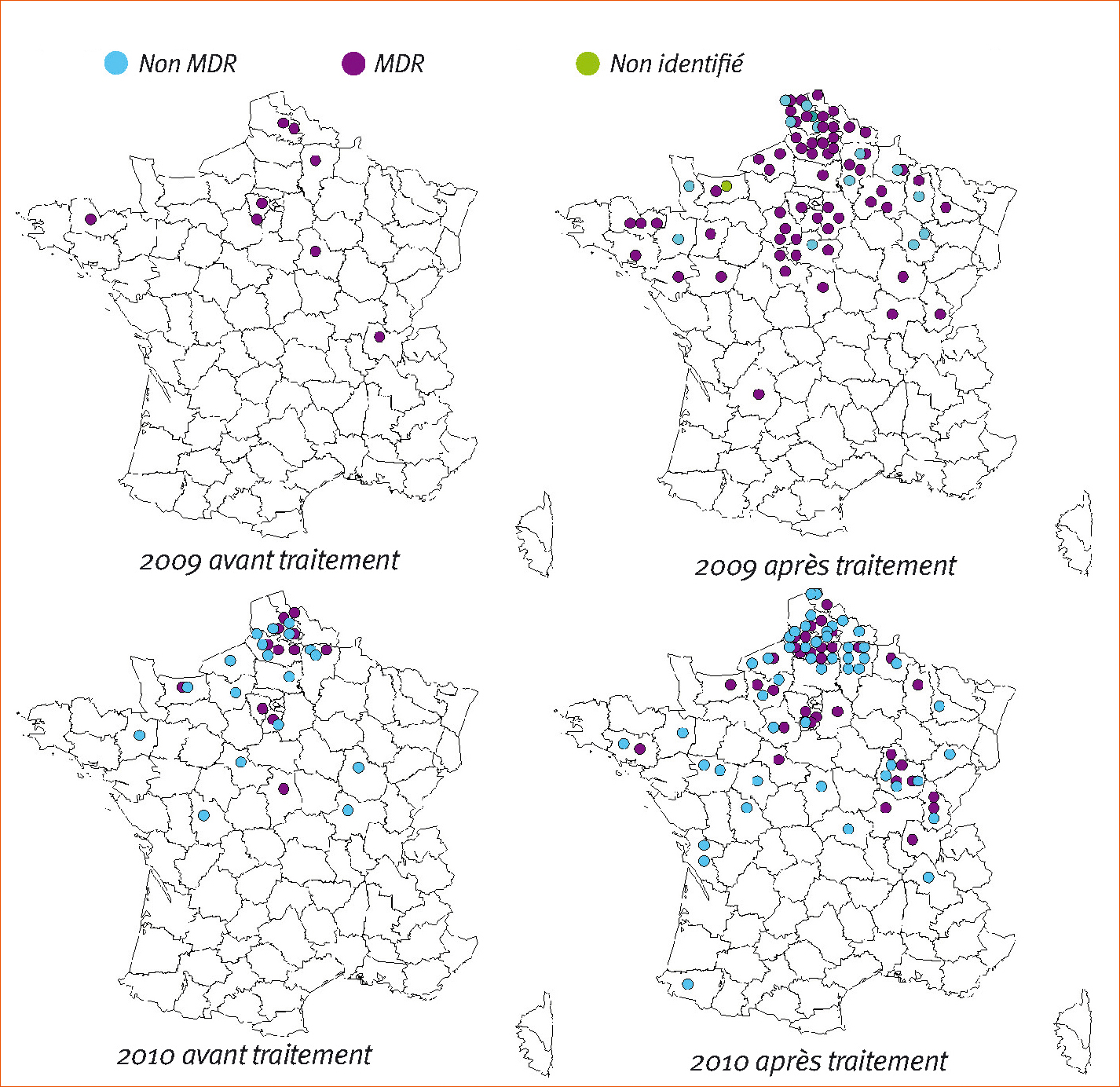
L'ensemble de ces nouveaux phénotypes est détecté depuis 2008 mais reste en fréquence faible dans les populations (Figure 1).
Ainsi en 2010, si ces souches sont présentes dans 30,3 % des populations analysées (20,7 % pour les souches « non MDR », soit TriR5+, TriR8+, TriR9 à TriR11, et 11,7 % pour l'ensemble des souches MDR), elles ne représentent en moyenne que 14,7 % des isolats dans ces parcelles (15,8 % pour les non MDR et 9,8 % pour les MDR).
Ces chiffres sont cependant en augmentation : les parcelles avec présence de souches émergentes représentaient 3 % de l'échantillonnage en 2008 puis 13 % en 2009.
Enfin, les cartographies (Figure 3) montrent que ces phénotypes résistants sont présents dans diverses régions françaises, avec une prépondérance pour le nord de la France. L'augmentation du nombre de sites positifs avant et après traitement suggère que ces phénotypes émergents sont bien sélectionnés par les fongicides. Enfin, la présence de ces isolats aux printemps 2009 et 2010, avant les premiers traitements, montre qu'au moins une partie de ces populations émergentes est suffisamment compétitive pour subsister pendant l'hiver.
À noter enfin que parmi ces souches émergentes, les non MDR sont largement détectées dans d'autres pays européens par les différents partenaires. Les souches MDR ont quant à elles été détectées en 2009 et en 2010 à la fois dans des populations françaises et britanniques.
Efficacité des fongicides
Aujourd'hui
La généralisation de souches TriMR, au début des années 2000, avait conduit à l'érosion de l'efficacité des triazoles. Ensuite cette efficacité a été renforcée par des associations entre IDM ou avec d'autres modes d'action. Depuis 2008, en dépit de l'émergence des nouveaux phénotypes en France, l'efficacité des programmes utilisant les meilleurs triazoles reste au moins identique aux années antérieures (Maumené et al., 2010), voire supérieure en cette année de faible pression parasitaire. Ce bon résultat semble contradictoire avec la situation décrite précédemment.
Certes, de nouvelles souches très résistantes viennent d'être détectées à des fréquences frisant parfois les seuils de détection des monitorings. Et c'est probablement parce que ces souches restent globalement rares et non généralisées qu'elles n'impactent pas l'efficacité des programmes fongicides. Il est difficile de prévoir si elles l'impacteront un jour.
Un essai en contamination artificielle mené à Boigneville (91) en 2010 a montré que la plupart des produits testés pouvaient être significativement (mais partiellement) affectés lorsque ces nouvelles souches sont dominantes dans la population.
Et demain
Potentiellement, et au vu des exemples malheureux rencontrés avec des isolats présentant des niveaux de résistance de cet ordre de grandeur (cas des QoI par exemple), l'efficacité des fongicides pourrait être compromise si, et seulement si, ces phénotypes nouveaux continuent d'augmenter en fréquence.
L'augmentation en fréquence de ces isolats dépend de nombreux facteurs, mais parmi eux, leur fitness (ou capacité à être compétitifs face aux autres souches ; ce paramètre reste à renseigner précisément) et la pression de sélection qui sera exercée sur les populations seront prépondérantes.
Il est donc heureux que ces phénotypes émergents aient été détectés et caractérisés de manière précoce sans avoir généré de résistance en pratique, car cela laisse l'opportunité de mettre en œuvre des stratégies anti-résistance permettant peut-être de ralentir leur évolution.
En particulier, puisque les souches dites MDR cumulent au moins deux mécanismes de résistance (altération de la cible et efflux accru) et présentent in vitro un spectre de résistance croisée entre les IDM, les QoI et les SDHI, il conviendrait d'estimer le pouvoir sélectif de ces différents modes d'actions sur les souches MDR. Le marché des fongicides céréales sera bientôt complété par plusieurs molécules SDHI et si la capacité des IDM à sélectionner les phénotypes émergents décrits plus haut semble acquise, il reste à confirmer l'impact sélectif que ce mode d'action nouvellement introduit pourra avoir, sur les souches MDR en particulier.
Plus globalement, il demeure important d'élaborer des recommandations limitant les diverses pressions de sélection et adaptées à ce contexte résistances en pleine évolution (Note nationale Céréales 2011).
Au final
Finalement, et malgré les difficultés techniques inhérentes aux années de faible pression parasitaire, 2010 a confirmé que les populations de S. tritici continuent d'évoluer. Les mécanismes de résistance aux IDM se sont diversifiés en même temps qu'ils se sont complexifiés.
Cela implique un effort accru en matière de recherche, tant au laboratoire qu'au champ.
Ceci rappelle également l'incroyable adaptabilité de ce champignon, déjà connu dans la littérature pour la variabilité génétique de ses populations et la grande plasticité de son génome. À ce titre, les prochaines campagnes seront également probablement très riches d'enseignements.
<p>* INRA, UR BIOGER-CPP, avenue Lucien-Brétignières. 78850 Thiverval-Grignon. walker@versailles.inra.fr</p>
Tableau 1 - Caractérisation in vitro des différents isolats de Septoria tritici présents dans les populations françaises pour leur résistance vis-à-vis des fongicides IDM. Les phénotypes émergents sont signalés en bleu. Les chiffres indiquent les niveaux de résistance moyens des phénotypes à un fongicide donné.
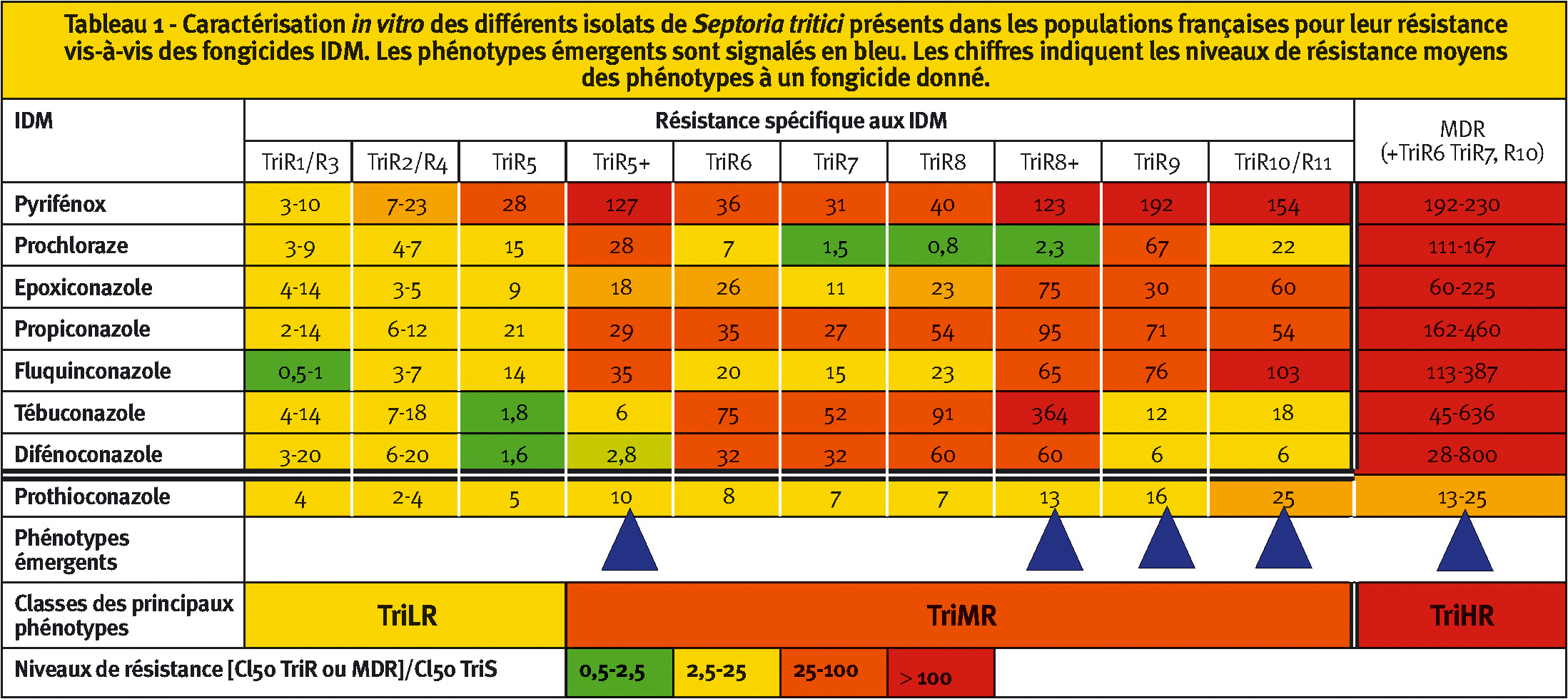
Figure 1 - Évolution de la résistance aux IDM dans les populations françaises de Septoria tritici. A - évolution de la structure des populations dans les modalités non traitées avant et après traitement en 2010. B - évolution au cours du temps des différentes catégories de souches résistantes aux IDM.
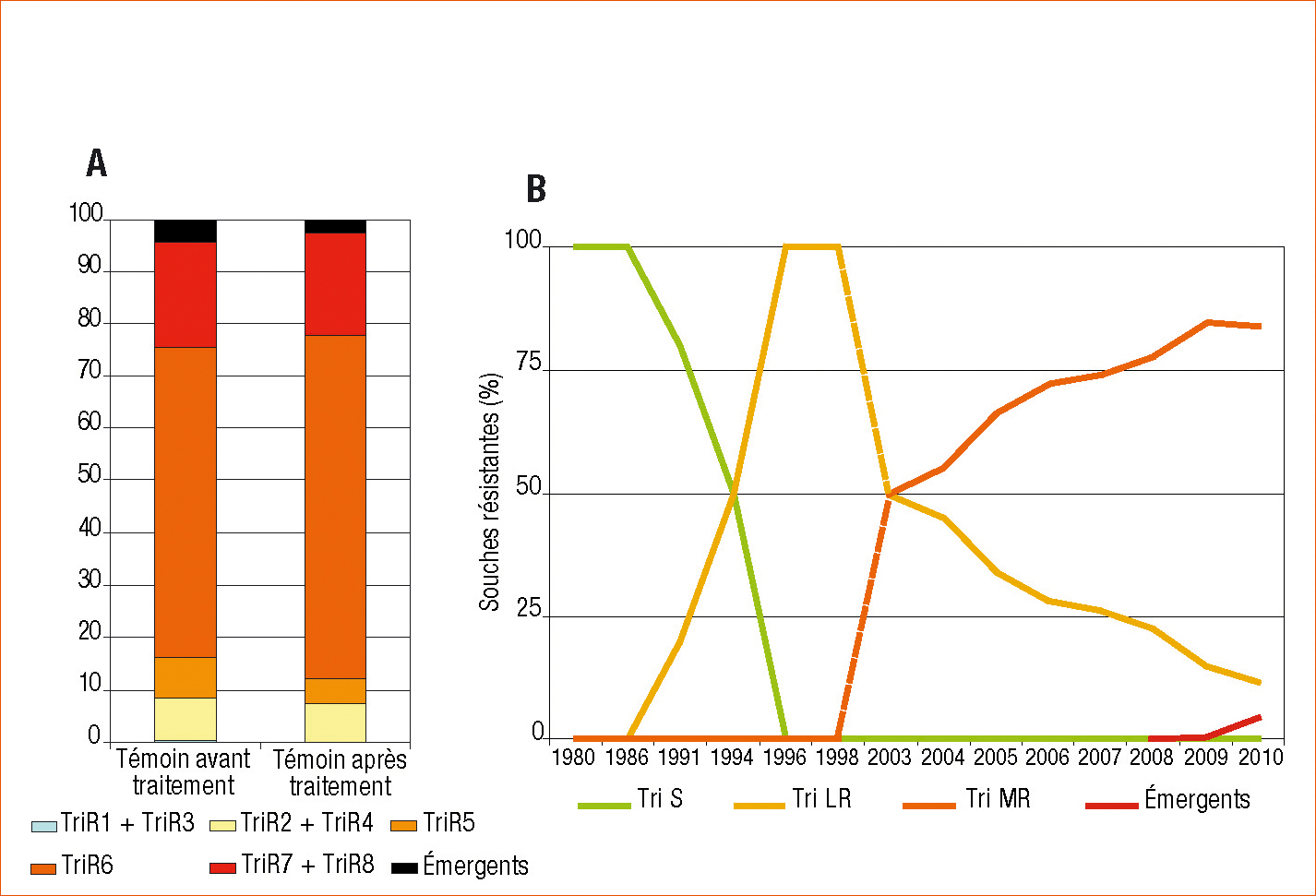
Figure 2 - Polymorphisme de la protéine cyp51, cible des triazoles, chez les différents isolats de Septoria tritici résistants aux IDM.
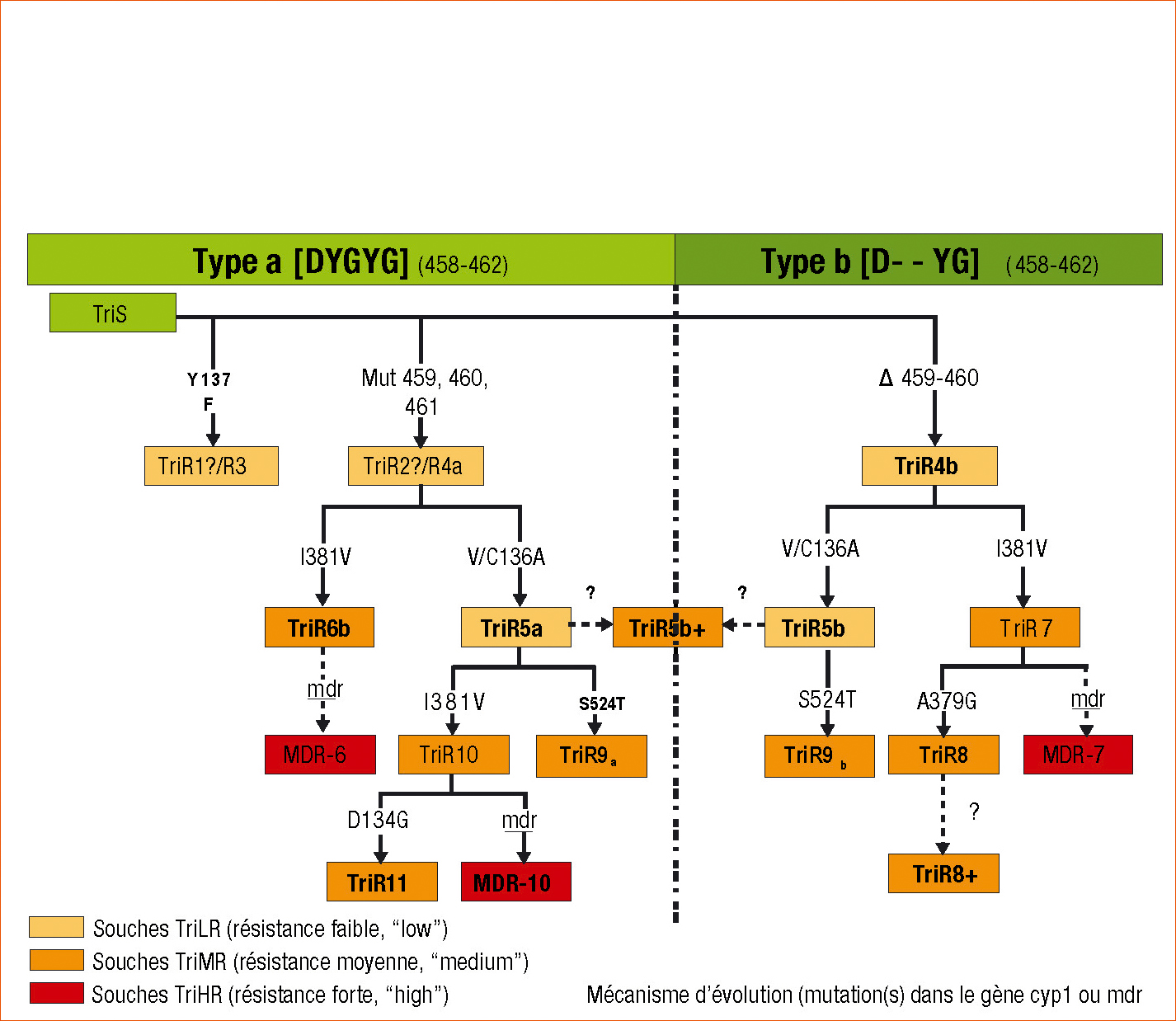
Cette présentation n'est pas un arbre phylogénétique. Elle ne montre pas la distance génétique entre souches. Mais elle relie chaque souche à celle dont elle dérive probablement, et suit la chronologie.
Par exemple les souches TRiR11 et MDR-10 ont évolué à partir de la souche TriR10, elle-même dérivant de la souche TRiR5a dont la souche TriR9a aussi « descend » probablement.
Figure 3 - Cartographie des sites présentant des phénotypes émergents et évolution entre 2009 et 2010.