 Imprimer
Imprimer
ph. Arvalis
 bien plus chargées en inoculum.")
Ci-dessus, résidus de colza. Dans notre étude, l'ADN de F. graminearum et de Microdochium sp. est quantifié début juin. Ces résidus pourraient donc être contaminants, mais moins que les cannes de maïs (médaillon en haut de page) bien plus chargées en inoculum.
, il reste moins de chaumes de blé à l'hectare que de cannes de maïs, donc moins d'inoculum prêt à infester le blé suivant. ... En fait le blé pourrait n'être pas toujours un précédent sans risque pour la fusariose à F. graminearum donc la contamination par le DON.")
Résidus de blé. En principe, un précédent blé est considéré comme moins porteur de « risque fusariose » qu'un précédent maïs. Mais, dans l'essai que nous avons réalisé, chaque gramme de résidu de blé contient au printemps un peu plus d'inoculum de fusariose qu'un gramme de résidu de maïs. Simplement, à façons culturales comparables (par ex. non-labour sans broyage, comme ci-contre), il reste moins de chaumes de blé à l'hectare que de cannes de maïs, donc moins d'inoculum prêt à infester le blé suivant. ... En fait le blé pourrait n'être pas toujours un précédent sans risque pour la fusariose à F. graminearum donc la contamination par le DON.
On sait que la fusariose de l'épi du blé est une maladie induite par plusieurs champignons des genres Fusarium et Microdochium, et que les Fusarium sp. sont capables de synthétiser des mycotoxines contaminant les grains. Ainsi F. graminearum peut produire du déoxynivalénol (DON). Cette fusariotoxine est réglementée et, selon l'ANSES(1), l'exposition de la population au DON par l'alimentation peut poser problème. Or l'occurrence de F. graminearum sur une récolte de blé dépend de divers facteurs, en particulier la présence et la nature des résidus de la récolte précédente. Mais il manque encore des données précises permettant d'évaluer le risque. L'étude présentée ici en apporte de nouvelles ; de plus, elle valide une technique analytique qui sera productive en données futures, à savoir la PCR en temps réel.
Sur le blé, la fusariose de l'épi est causée par plusieurs espèces de champignons, en majorité Fusarium graminearum et Microdochium spp. Outre le risque de perte de rendement, la maladie peut s'accompagner de l'accumulation dans les grains de mycotoxines, dont le déoxynivalénol (DON). Cette toxine est réglementée en Europe depuis 2006 pour les blés destinés à l'alimentation humaine. Ainsi, le respect des limites réglementaires est devenu une condition d'accès au marché. De plus l'EAT, « Étude de l'alimentation totale » dont les résultats ont été publiés en juin 2011 par l'ANSES(2), a montré que l'exposition alimentaire au DON, venant surtout de produits céréaliers, représente un « risque ne pouvant être écarté pour certains groupes de consommateurs ». Le DON est un défimajeur pour les filières céréalières.
La question du précédent cultural et de ses résidus
Un rôle reconnu... et des inconnues
De nombreuses études ont mis en évidence l'impact des pratiques culturales et des facteurs environnementaux sur le développement de F. graminearum et la production de DON.
Parmi les facteurs de risque identifiés, le précédent cultural ressort comme un des principaux. Mais le classement des précédents par rapport au risque mycotoxines associé n'est pas le même partout. Ainsi, un précédent blé tendre est classé à risque faible en France et au Royaume-Uni, à risque moyen en Allemagne et en Suède, et à risque fort en Italie. Ces différences posent la question du rôle des résidus dans l'épidémiologie de la maladie.
La présence dans le sol de résidus de la récolte précédente est décrite dans la littérature comme déterminante, les Fusarium et Microdochium étant capables de survie saprophyte. Cette aptitude est particulièrement marquée chez F. graminearum (Wilcoxson et al., 1988 ; Naef and Defago, 2006). Elle reste peu documentée sur Microdochium spp.
Demain, davantage de résidus des cultures précédentes ?
Ainsi, les résidus de culture subsistant au sol constituent la source majeure d'inoculum primaire susceptible d'infecter la culture suivante (Luongo et al., 2005 ; Bateman et al., 2007 ; Caron et al., 2007). Or, le contexte à venir de réduction de 50 % des pesticides dictés par le plan Ecophyto 2018, associé à une augmentation du non-labour dans les exploitations, pourrait favoriser durablement la persistance de ces niches refuges pour l'inoculum primaire.
La caractérisation des résidus de culture quant à leur rôle dans l'épidémiologie de la maladie devient donc une composante particulièrement importante à étudier afin de proposer des outils de gestion adaptés visant à réduire les risques d'infection.
Des études existaient, mais...
Certaines études ont poussé les recherches jusqu'au suivi du potentiel infectieux sur différents types de résidus. Mais leurs méthodologies étaient très variables et leurs résultats parfois contradictoires (Khonga et Sutton, 1988 ; Dill-Macky et Jones, 2000 ; Caron et al., 2007 ; Köhl et al., 2007 ; Peyreira et al., 2008 ; Güo et al., 2010 ; Hogg et al., 2010).
Or le développement récent des méthodes (dites PCR en temps réel) de quantification de l'ADN sur des matrices diversifiées permet d'étudier de manière plus fine et plus rapide les espèces fongiques présentes sur ces résidus (Güo et al., 2010 ; Hogg et al., 2010).
La PCR, outil pour mieux comprendre
C'est pourquoi une étude de caractérisation de différents types de résidus a été entreprise à l'aide de cette méthode sur la station expérimentale de Boigneville. Couplée à ce dispositif, une enquête avec prélèvements de résidus dans les parcelles d'agriculteurs a été réalisée en collaboration avec le Comité technique blé dur de la région Centre d'une part et Bayer CropScience d'autre part.
L'objectif de cette étude était d'une part de valider la méthode d'analyse des espèces ciblées F. graminearum et Microdochium spp. par PCR en temps réel, d'autre part d'évaluer dans différents types de résidus en dynamique, ou lors de la floraison, leur potentiel infectieux.
Deux voies d'étude complémentaires
Suivi dynamique : 4 dates de prélèvements à Boigneville
Dans cette étude, des échantillons de résidus de betterave, colza, tournesol, blé et maïs récoltés en 2009 sont prélevés en continu sur 5 diagonales des parcelles identifiées. Les prélèvements sont réalisés juste à la récolte des cultures, puis en décembre 2009, avril 2010 et enfin début juin, à la floraison des blés. L'objectif de ce dispositif était d'évaluer la cinétique de F. graminearum dans le temps.
Sondage sur 30 parcelles en région Centre
Pour compléter l'étude réalisée sur la station de Boigneville et afin de multiplier les conditions pédo-climatiques, un sondage est réalisé dans la région Centre suivant la même méthodologie. Les prélèvements sont effectués uniquement lors de la floraison des blés (juin 2010).
L'objectif de ce dispositif est de faire un premier sondage en démultipliant les conditions pédo-climatiques et de suivre spécifiquement F. graminearum et Microdochium spp.
Gestion des échantillons
Les échantillons récoltés sont séchés dans des conditionneurs à une température n'excédant pas 60 °C pour éviter la dénaturation de l'ADN tout en stoppant l'évolution des champignons. Une fois secs, ces échantillons sont broyés, d'abord grossièrement via un broyeur à couteaux avec une grille de diamètre 4 mm, puis avec un broyeur à godets de 35 ml avec bille pendant 30 s. Les échantillons broyés sont conservés à + 4 °C jusqu'à l'analyse.
Extraction de l'ADN
50 mg de broyat de chaque échantillon sont aliquotés dans une plaque 96 puits. L'ADN total est extrait via un kit (Macherey-Nagel) puis dosé au spectrophotomètre et dilué à 10 ng/µl. La dilution est validée par un nouveau dosage au spectrophotomètre. Les ADN extraits sont conservés à – 20 °C en attente de l'amplification.
PCR en temps réel
Les quantifications de F. graminearum, Microdochium nivale et Microdochium majus sont réalisées en utilisant la technologie Taqman. Les amorces spécifiques à chaque espèce fongique recherchée permettent l'amplification de l'ADN du pathogène. Sa quantification est rendue possible grâce à l'utilisation d'une gamme standard réalisée avec de l'ADN de champignon pur. Les données brutes d'ADN de champignon obtenues sont ensuite normalisées par rapport à 100 ng d'ADN total. Enfin, les résultats sont exprimés en logarithme de la quantité d'ADN fongique par rapport à la quantité d'ADN total (végétal et champignon) de l'échantillon.
Résultats
Sondage : les deux espèces partout, mais des taux différents
La figure 1 présente les résultats du sondage en région Centre. Ce premier sondage permet de faire plusieurs constats.
D'abord, les deux espèces fongiques sont quantifiées sur les 5 résidus de culture étudiés.
Néanmoins la quantité d'ADN de F. graminearum est plus forte dans les résidus de betterave, blé et maïs comparativement aux résidus de colza et tournesol. Ce résultat peut s'expliquer par le fait que F. graminearum est un pathogène connu de la betterave, du blé et du maïs et non du colza et du tournesol. Néanmoins, même à des niveaux faibles, il semblerait que les cannes de colza et de tournesol puissent être colonisées par ce champignon.
Concernant Microdochium, la seule tendance nette est une quantité d'ADN un peu plus élevée dans les résidus de maïs.
Maïs et blé sont les plus infestés
En conclusion, et malgré le faible jeu de données, ce sondage permet de valider la méthode de PCR en temps réel utilisée sur les différentes matrices. Même si les niveaux d'infection sont difficiles à évaluer en valeur absolue, il apparaît en tendance que la population fongique est différente selon le type de résidus. De plus, on confirme que le blé et le maïs sont les résidus les plus infectés par F. graminearum.
Ces résultats préliminaires sont assez bien corrélés avec ceux de la littérature (Peyrera et al., 2008 ; Köhl et al., 2007).
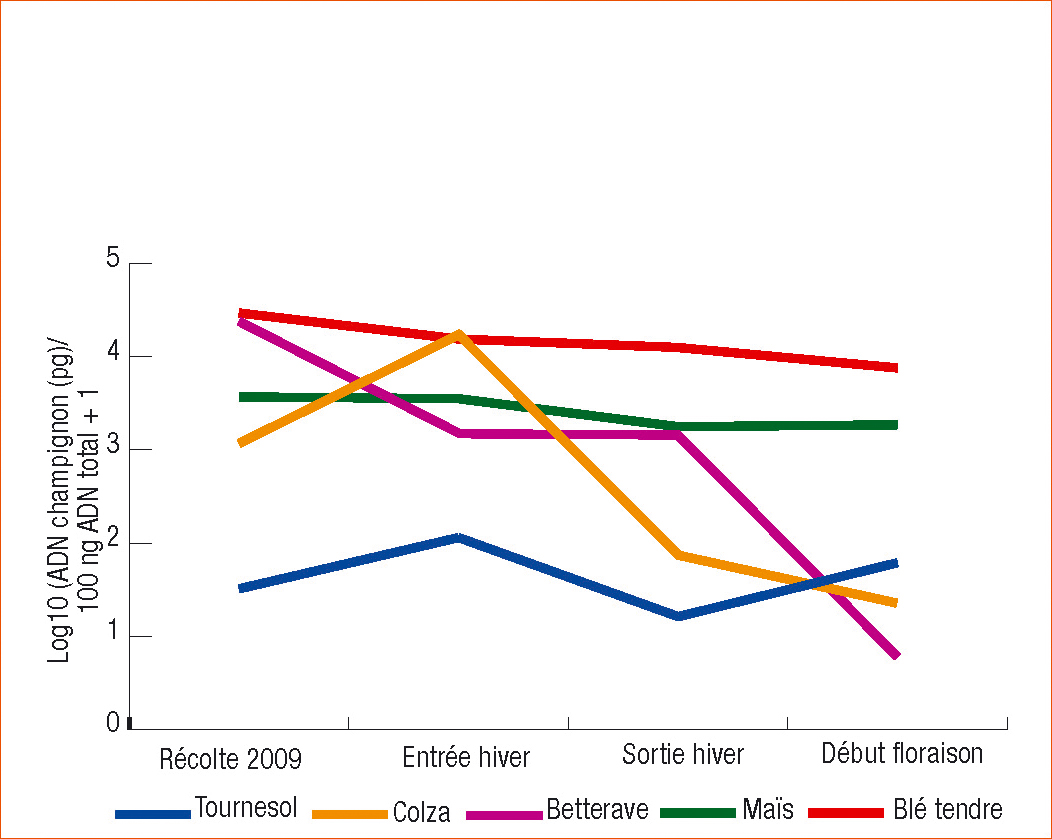
Suivi dynamique
Les résultats du suivi dynamique sont reportés en figure 2. Deux types de cinétique se distinguent.
Il ne semble pas y avoir d'évolution dans le temps sur les résidus de maïs, de blé et de tournesol. Mais ce dernier reste toujours nettement moins infecté que les céréales.
En revanche la quantité de champignon détectée diminue dans le temps sur les débris de colza et surtout de betterave. Ces derniers, aussi contaminés que ceux du blé à la récolte (la betterave est sensible à la fusariose), sont les moins contaminés à la floraison du blé suivant, peut-être en lien avec une décomposition des débris plus rapide.
Il n'y a aucun cas où la quantité de champignon augmente dans le temps ; ainsi l'hypothèse d'une colonisation des résidus à la sortie de l'hiver n'est pas validée par cette étude.
Les résultats observés sur les prélèvements effectués lors de la floraison des blés confirment les résultats précédents à l'exception de la betterave.
À ce moment-là, deux groupes de précédents se distinguent : d'un côté le blé et le maïs, les plus fortement infectés et, de l'autre, le tournesol, le colza et la betterave.
Attention, c'est « au gramme de résidu » !
Néanmoins, ces résultats sont à pondérer par la quantité de résidus présente sur la parcelle à cette période. En effet, les résultats sont exprimés par gramme de résidus. Or en semis direct, la quantité de maïs au sol est nettement supérieure à celle d'un blé par exemple. La quantité d'inoculum sera donc elle aussi supérieure.
Le travail continue
Cette première étude confirme l'impact de la gestion des résidus sur la pression de l'inoculum. Elle offre une piste pour comprendre l'apparition de fortes teneurs en toxines dans des parcelles à rotation ne présentant a priori pas de risque agronomique particulier.
La validation de l'utilisation de l'outil PCR en temps réel pour l'étude des résidus permet de poursuivre le travail de manière plus approfondie en 2011. Cela devrait conduire à une meilleure caractérisation des résidus de culture quant à leur potentiel infectieux par rapport à F. graminearum et Microdochium spp, en lien avec leur composition biochimique notamment le rapport carbone/azote, leur capacité à être dégradé et le climat, composante indispensable à la compréhension de la maladie et non travaillé dans la présente étude.
Cette nouvelle expérimentation nous amènera à étudier également les espèces présentes sur les grains ainsi que les teneurs en mycotoxines permettant ainsi d'établir les liens, s'ils existent, entre potentiel infectieux sur résidus, quantité de champignon sur grain et toxines produites en lien avec le climat.
<p>* Arvalis-Institut du végétal, 91720 Boigneville.</p> <p>(1) Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail.</p> <p>(2) Avis de l'Anses, saisine n° 2006-SA-0361, EAT (Étude de l'alimentation totale) 2, 2006-2010. Voir aussi p. 13 de ce numéro.</p>
Figure 1 - Quantification moyenne de F. graminearum et Microdochium spp sur 5 types de résidus (tournesol n=10, colza n=7, betterave n=2, blé n=4, maïs n=7) prélevés lors de la floraison des blés en région Centre. Résultats exprimés par gramme de résidus.
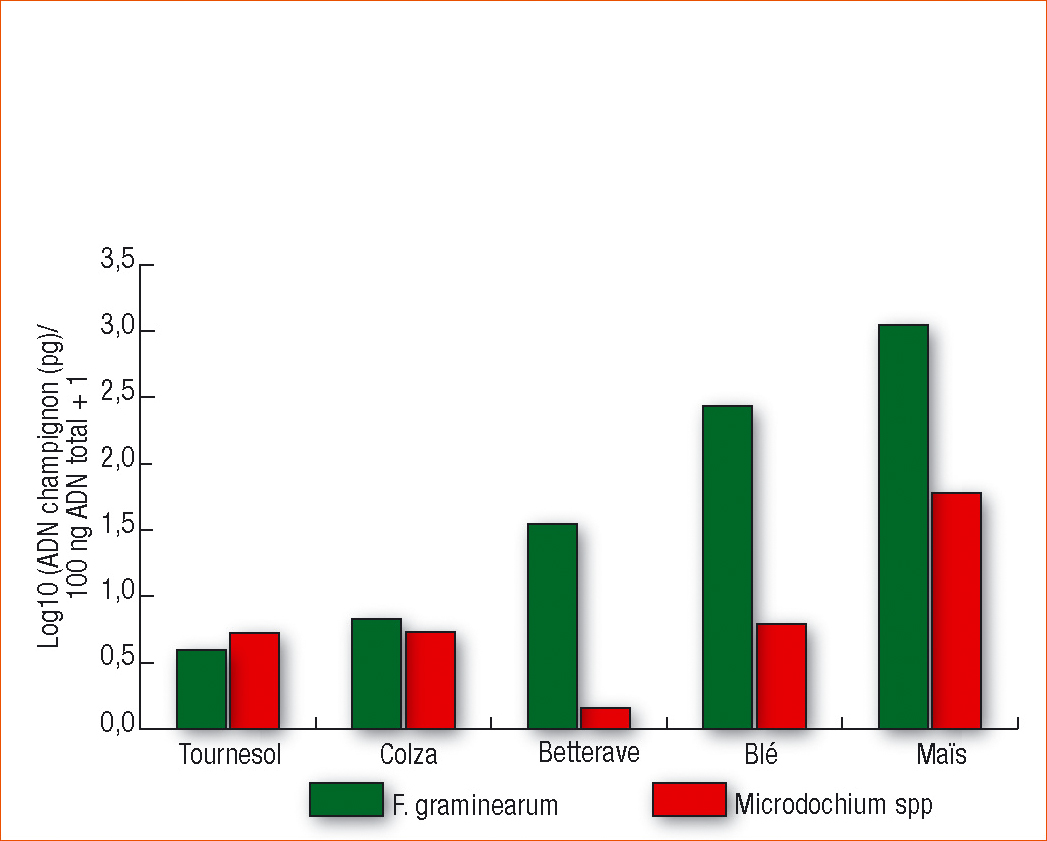
Figure 2 - Quantification de F. graminearum sur 5 différents types de résidus (tournesol, colza, betterave, blé, maïs) prélevés à différentes période de l'année, lors de leur récolte, en décembre (entrée hiver), en avril (sortie hiver) et en juin (début floraison). Résultats exprimés par gramme de résidus.
, qui peut produire du DON.")

