 Imprimer
Imprimer sur blé. Pascal Boulin, BayerCropScience")
Symptôme de pied chétif (WDV) sur blé. Pascal Boulin, BayerCropScience
Aujourd'hui, le principal moyen de protéger le blé et l'orge du WDV est de combattre les insectes vecteurs du virus responsable du « pied chétif ». Certes. Mais on connaît encore mal le « fonctionnement de ce pathosystème » : notamment les facteurs favorisant l'infection des plantes par le virus. Donc le risque de traitement non justifié existe. Cet article fait le point sur la question, avec deux objectifs :
– présenter l'état des lieux des connaissances sur les partenaires impliqués dans la maladie du « pied chétif » ;
– envisager les orientations de recherche à privilégier pour progresser dans la compréhension de la dynamique d'extension des épidémies de WDV et améliorer la qualité sanitaire des céréales tout en raisonnant les interventions chimiques.
Le « pied chétif », pourquoi l'étudier
Une maladie transmise par des insectes
Comme toutes les plantes, les céréales sont sensibles à divers pathogènes : champignons, bactéries, nématodes et virus. Parmi les bioagresseurs viraux, le virus du pied chétif du blé (Wheat dwarf virus, WDV) occupe une place importante dans la hiérarchie des problèmes sanitaires liés à ces cultures.
Ce statut résulte en partie du fait que les moyens de lutte disponibles contre ce phytopathogène sont extrêmement limités. En effet, l'absence de sources de résistance efficace au WDV chez le blé ou l'orge [malgré quelques sources de tolérance chez l'orge récemment décrites en Hongrie (Benkovics et al., 2010)], associée à l'inexistence de produits capables d'agir directement sur les virus, conduit les agriculteurs à mettre en place des méthodes de lutte indirectes pour protéger leurs cultures. Ainsi, le principal moyen de lutte contre le WDV est l'utilisation d'insecticides en traitement de semences et/ou en végétation.
En effet, ce phytovirus est transmis de plantes à plantes exclusivement par l'intermédiaire d'un insecte : des cicadelles du genre Psammotettix. L'élimination des insectes dans les cultures de céréales permet de prévenir les infections potentielles. Cependant, l'utilisation de produits phytosanitaires engendre des coûts directs (prix du produit et de ses applications) et indirects (coût potentiel pour l'environnement et sélection possible d'insectes résistants) à prendre en compte dans la mise en place de stratégies de lutte durable et raisonnée.
Mieux connaître pour ne traiter qu'en cas de besoin
Compte tenu des éléments dont disposent les agriculteurs pour appréhender la problématique « pied chétif », des décisions d'intervention risquent d'être prises alors qu'un risque viral n'est pas avéré.
Aussi, pour optimiser l'utilisation des insecticides, il est important d'identifier et de hiérarchiser les paramètres biotiques (espèces d'insectes vecteurs, variabilités du virus, sensibilité des plantes hôtes, effet du paysage, etc.) et abiotiques (paramètres environnementaux) impliqués dans l'incidence de la maladie sur céréales.
Cet objectif nécessite de disposer de connaissances sur le fonctionnement du pathosystème concerné. Or, les données disponibles à ce jour sur le WDV et sur les interactions WDV/vecteurs/hôtes sont encore limitées. Il est donc indispensable de progresser dans l'étude de ce pathosystème pour, à terme, nous permettre de raisonner les interventions et d'adapter les pratiques culturales en fonction des risques épidémiques réels.
Ce qu'on sait de la maladie
Caractéristiques générales de la maladie
Le « pied chétif » est une des maladies virales les plus dommageables sur céréales. Principalement décrit sur orge et blé, il est présent en Europe, Asie et Afrique. Compte tenu des altérations (Vacke J., 1972) provoquées par le WDV sur ses hôtes (nanisme, jaunisse et, dans certains cas, mort de la plante), les épidémies de « pied chétif » peuvent entraîner des pertes de rendement allant jusqu'à 75 % (Fohrer et al., 1992).
Malgré ces impacts économiques et agronomiques majeurs, les connaissances scientifiques relatives à l'épidémiologie de cette maladie restent limitées. Depuis la première description du WDV en 1961 dans l'ancienne Tchécoslovaquie (Vacke, 1961) et jusqu'à la fin des années 1980, peu d'études ont été conduites sur la maladie. Mais le constat sanitaire défavorable décrit sur céréales depuis deux décennies a relancé l'intérêt de la communauté scientifique et de la filière. En effet, après une période de moindre importance agronomique (seconde moitié du XXe siècle), ce phytopathogène a ré-émergé à la fin des années 1990 dans l'Est et le Nord de l'Europe (ex. : Hongrie, Suède, Allemagne et Finlande), au Moyen Orient (ex. : Turquie), en Asie (ex. : Chine) et en Afrique (ex. : Tunisie et Zambie).
Depuis 2003, les foyers français du quart nord-est se propagent vers le centre et l'ouest (Fig. 1). Cette zone d'activité du WDV correspond à une zone géographique d'infestation des cicadelles du genre Psammotettix. Ces données sont cohérentes avec les connaissances publiées sur la transmission du WDV par ces cicadelles.
Infections primaires, secondaires, stade de sensibilité des plantes
Le cycle de la maladie sur céréales débute à l'automne, lorsque les cicadelles virulifères (porteuses de virus) migrent de leurs réservoirs estivaux (graminées sauvages) vers les parcelles de céréales (Fig. 2).
Toutefois, des données collectées en Suède montrent que l'impact sanitaire des réservoirs sauvages se limite aux infections primaires des parcelles de céréales (5 % des plantes de la parcelle, d'après Lindblad et Sigvald, 2004). L'extension des foyers infectieux au sein des parcelles repose principalement sur des infections secondaires intra-parcellaires.
Cependant, le rôle respectif des différents paramètres impactant l'émergence et l'extension de ces foyers infectieux n'a pas été clairement défini. Quelques éléments sont néanmoins documentés.
Ainsi, au stade « jeune plante », le blé et l'orge sont particulièrement sensibles à l'infection par le WDV. Cette sensibilité se maintient jusqu'à la montaison (stade 31 ; premier nœud 1 cm au-dessus du plateau de tallage). Ensuite, une résistance de maturité de l'hôte vis-à-vis du WDV apparaît.
A propos du vecteur
Les espèces du genre Psammotettix
Les cicadelles du genre Psammotettix (Hemiptera, Cicadellidae, Deltocephalinae) sont des insectes holarctiques, piqueurs-suceurs, communément retrouvés dans les parcelles de céréales, sur Poacées sauvages et dans les zones de repousses de céréales (Lindblad et Areno, 2002).
Plusieurs critères morphologiques permettent d'identifier les cicadelles du genre Psammotettix (forme de la tête, taches sur l'écusson, longueur et pigmentation des ailes ; Vilbaste, 1982 ; Della Giustina, 1989). Mais l'accès à l'espèce passe par la dissection et l'observation de l'édéage, organe reproducteur mâle (Biedermann et Niedringhaus, 2004). Il est impossible de déterminer l'espèce des cicadelles femelles du genre Psammotettix à l'aide de clés de détermination classiques.
La littérature décrit 53 espèces en Europe dont 18 présentes en France. Cependant, des incohérences (confusion d'espèces, synonymies entre espèces) présentes dans la littérature, reflètent la difficulté d'identification spécifique basée sur l'édéage. Afin de clarifier la situation, une approche visant à définir les espèces de Psammotettix sur des critères moléculaires doit être entreprise (Le Roux et Rubinoff, 2009).
P. alienus vecteur principal ?
Selon la plupart des données bibliographiques, Psammotettix alienus (Dahlb.) serait le vecteur principal de WDV (ex., Zhang et al., 2010). Cependant, l'identité spécifique des insectes utilisés dans le cadre de ces études n'est que très peu documentée dans les articles correspondants, ce qui :
– augmente l'incertitude liée à l'identité spécifique des cicadelles manipulées ;
– pose question sur l'efficacité de vection du WDV par chaque espèce de Psammotettix.
Ainsi, on trouve dans la littérature des données indiquant que le vecteur principal de WDV serait Psammotettix provincialis (Ekzayez et Kumari, 2011), sans éléments factuels à l'appui de cette affirmation.
Dans l'hypothèse où les différentes cicadelles Psammotettix n'auraient pas la même efficacité de transmission, ce paramètre serait particulièrement important pour appréhender l'épidémiologie du WDV.
Cycle biologique de P. alienus
Le cycle biologique de la cicadelle P. alienus (Dahlb.) a été étudié en détail par Manurung et al. (2005) et Guglielmino et Virla (1997). Il débute par le dépôt d'un œuf dans les couches superficielles d'une feuille de céréale (Fig. 2). L'intensité de ponte dépend des conditions environnementales (températures) et de l'hôte (espèce végétale) sur lequel pond la cicadelle. L'œuf passe l'hiver sur la plante et débute son développement en sortie d'hiver avec l'augmentation des températures et de la longueur des jours. Ce développement dure en moyenne 18 jours (à 20 °C, 70-95 % RH, 18 heures de lumière par jour). A terme, l'éclosion permet de produire une larve de premier stade (L1) (Fig. 2).
Cette larve passe par cinq stades (de L1 à L5). En conditions de cultures optimales (cf. ci-dessus), la durée totale du développement larvaire est en moyenne de 32 jours. La dernière étape du développement correspond au stade adulte (mâle et femelle) pendant lequel la cicadelle produit par reproduction sexuée de nouveaux œufs. Le cycle biologique (de l'œuf à l'adulte) de cet insecte dure environ 51 jours. La première ponte s'effectuant à partir du 7e jour du stade adulte, la durée du cycle œuf-œuf est en moyenne de 58 jours.
Diverses études sur la dynamique de populations des Psammotettix ont montré que :
– la densité d'individus peut atteindre 43 cicadelles adultes/m² (Manurung et al., 2005),
– le sex-ratio des populations présentes dans les parcelles est proche de l'équilibre (mâles/femelles ≈ 1),
– les conditions climatiques du nord de la France permettent aux cicadelles de réaliser 4 cycles du printemps à l'automne.
Tous les stades de développement sont compatibles avec la transmission virale. Les larves sont beaucoup plus mobiles que les adultes, mais ces derniers se déplacent à plus longue distance grâce à leurs ailes.
Enfin, une analyse de l'interaction cicadelles/autres ravageurs a montré que la présence de pucerons sur une plante modifie le comportement de la cicadelle en la perturbant dans ses phases d'installation, ce qui la conduit à visiter plus de plantes (Alla et al., 2001). Cette caractéristique pourrait impacter la dynamique d'extension des foyers infectieux.
L'agent pathogène
Caractéristique du WDV
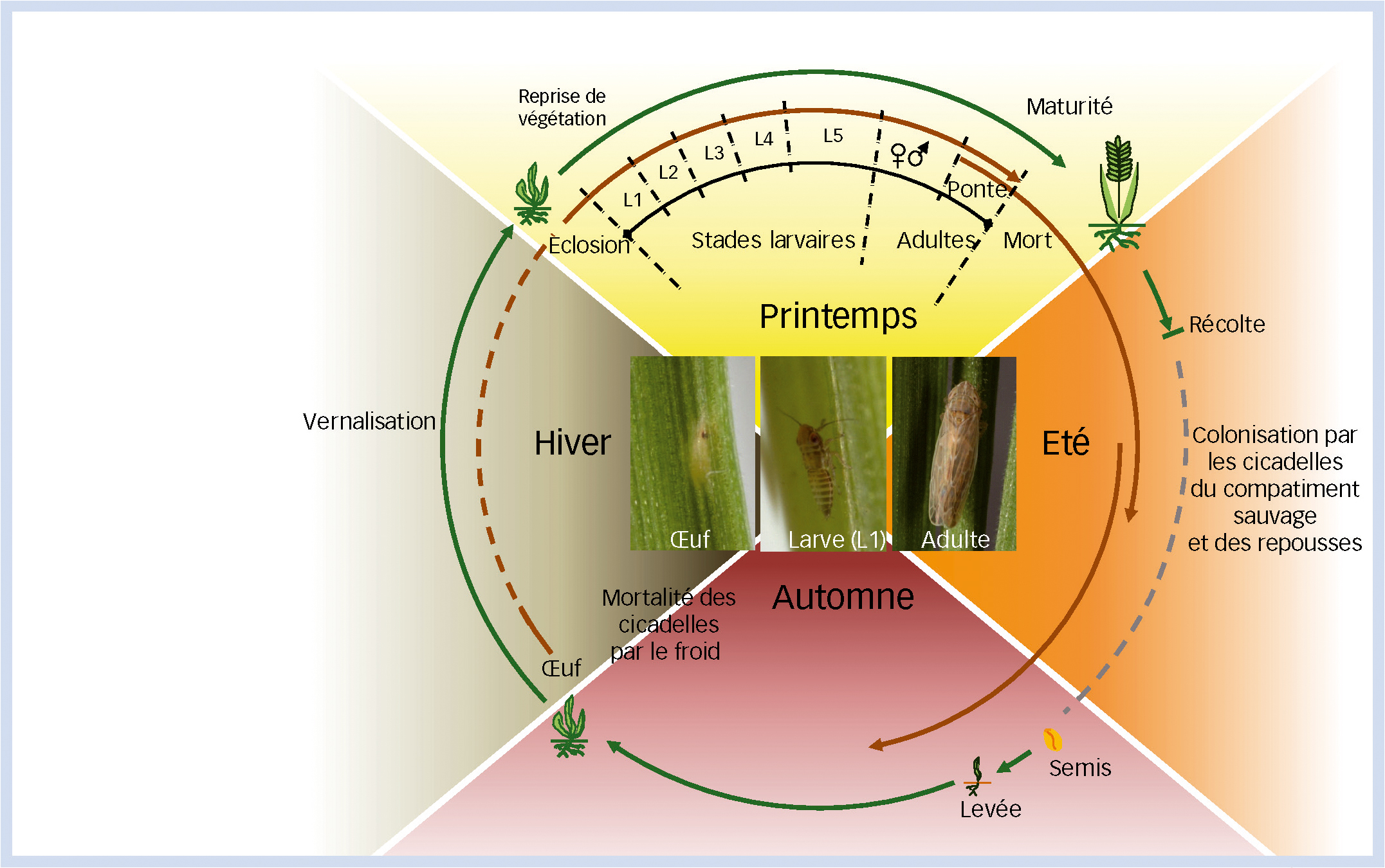
Le virus du pied chétif du blé (famille Geminiviridae, genre Mastrevirus), possède une capside en double icosaèdre de 30 nm de long sur 18 nm de large (Fig. 3).
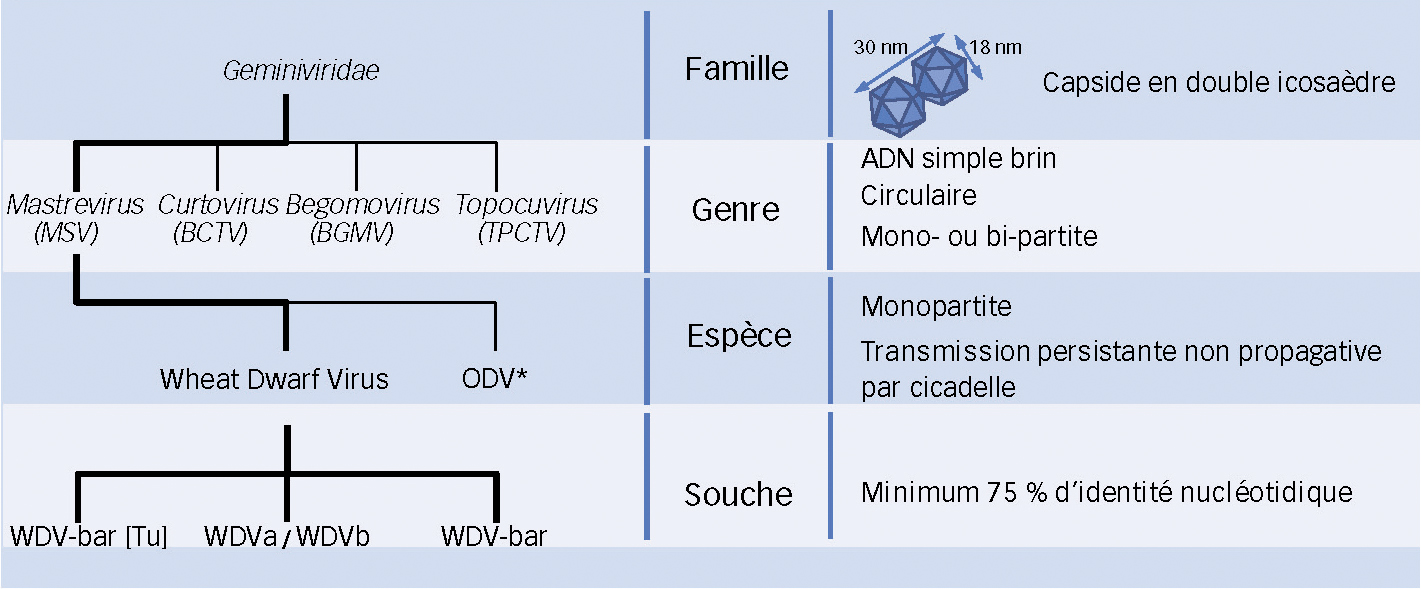
Le génome viral monopartite (Fig. 4) est constitué d'une molécule d'ADN simple brin circulaire de 2 749 bases codant pour trois protéines fonctionnelles correspondant à la polymérase (constituée de deux protéines Rep/RepA dont le rôle est d'assurer la réplication du génome viral), à la protéine de capside (formant la particule virale mature) et à la protéine de mouvement (impliquée dans la migration du virus dans la plante hôte). Le génome présente deux régions intergéniques (LIR et SIR) importantes pour la réplication et l'expression des protéines (Kvarnheden et al., 2002).
Transmission : mode persistant non propagatif
Le WDV est transmis par des cicadelles du genre Psammotettix selon le mode persistant non-propagatif (pour une revue, Brault et al., 2010). Ainsi, il ne se multiplie pas dans la cicadelle et n'est pas transmis de manière verticale à la descendance de l'insecte.
À l'éclosion, la larve commence à s'alimenter sur les plantes environnantes. En arrivant sur une plante infectée, la larve se charge en particules virales lors de la prise alimentaire et devient virulifère dès lors que la période de latence (quelques heures minimum) requise par le virus pour traverser les différents tissus/organes de l'insecte (intestin, hémolymphe et glandes salivaires) est achevée.
À partir de ce stade, les prises alimentaires suivantes permettront l'inoculation de particules virales à de nouvelles plantes.
Les larves restent à l'état virulifère pendant plusieurs jours/semaines. Ce stade virulifère n'est pas modifié par les mues successives. Enfin, compte tenu de l'importante mobilité de ces insectes, une très faible densité de cicadelles virulifères peut potentiellement permettre l'inoculation de virus à un nombre important de plantes.
Gamme d'hôtes et variants du virus
La gamme d'hôtes du WDV inclut le blé, l'orge, l'avoine et de nombreuses graminées sauvages (Schubert et al., 2007). Les caractéristiques moléculaires des isolats de WDV présents dans les graminées sauvages sont semblables à celles du WDV présent sur les surfaces cultivées. Toutefois, il semble exister dans ces deux compartiments deux variants de WDV (WDVa et WDVb) dont la différence génétique n'est, en moyenne, que de 1,4 % (Ramsell et al., 2008).
Récemment, des souches virales différentes du WDV décrit précédemment, WDV-bar (Lindsten et Vacke, 1991) et WDV-bar[Tu] (Köklü et al., 2007), ont été décrites sur la base de leurs caractéristiques moléculaires (variabilité de séquence) et/ou biologiques (gamme d'hôtes différentielle). Les souches WDV et WDV-bar diffèrent par des variations moléculaires de l'ordre de 16 % alors que les isolats de souche WDV présentent entre eux moins de 3 % de variabilité (Kvarnheden et al., 2002).
L'analyse des caractéristiques moléculaires de l'isolat WDV-bar[Tu], isolé sur orge en Turquie, semble montrer que ce dernier est issu de la recombinaison entre un isolat de type WDV-bar et un virus du genre Mastrevirus encore inconnu (Ramsell et al., 2009). Les souches WDV et WDV-bar possèdent une gamme d'hôtes similaire (Lindsten et Vacke, 1991).
Toutefois, le WDV-bar est principalement inféodé à l'orge (« bar »/« barley » ; orge en anglais) alors que les souches WDV sont décrites à la fois sur orge et sur blé (Kundu et al., 2009).
Durant des années, les données acquises sur l'épidémiologie du pied chétif semblaient indiquer que les souches WDVbar n'infectaient pas le blé. Cependant, une étude récente visant à décrire, sous conditions contrôlées, la capacité des souches de WDV à infecter différentes Poacées a montré qu'une souche WDV-bar peut infecter le blé (Ramsell et al., 2009). Ce résultat obtenu au laboratoire a été confirmé par la description d'un isolat WDV-bar (isolat Odessa) dans une parcelle de blé en Hongrie (Tobias et al., 2011).
En fait, il semblerait que lorsqu'ils infectent le blé, les isolats de WDV-bar soient plus agressifs que les isolats WDV.
Découverte de l'ODV, « O » comme Oat, l'avoine
L'existence de ces différentes souches de WDV complexifie significativement l'épidémiologie de cette maladie. Elle exige de prendre en compte ces diverses espèces/variants viraux comme plusieurs agents pathogènes responsables, seuls ou en association à l'échelle de la parcelle ou d'une région, de symptômes de « pied chétif ».
De plus, une nouvelle espèce virale, l'Oat dwarf virus (ODV) proche du WDV (voir Fig. 3), a été décrite comme agent causal de « pied chétif » sur avoine (Schubert et al., 2007). Ceci ajoute un niveau de complexité supplémentaire dans l'étude de cette maladie sur céréales.
Techniques de diagnostic pour identifier espèces et variants
En complément du diagnostic visuel, basé sur l'observation des symptômes de l'infection virale, différentes techniques sont disponibles pour valider la présence des souches/variants de WDV dans des vecteurs et/ou du matériel végétal.
Compte tenu des caractéristiques des constituants viraux (capside protéique et acide nucléique, voir fig. 3) des différentes souches de WDV, ces dernières peuvent être distinguées à l'aide d'approches sérologiques (ELISA, Vacke et Cibulka, 2000) et moléculaires [PCR (Commandeur et Huth, 1999) ; RCA-RFLP (Schubert et al., 2007) et PCR quantitative (Zhang et al., 2010)].
La spécificité des tests disponibles va de l'espèce-spécifique (détection de tous les WDV, pour exemple : anticorps polyclonaux) à la souche-spécifique [détection spécifique d'une souche virale, pour exemple : PCR quantitatives spécifiques de WDV ou de WDV-bar (Gadiou et al., 2011)].
L'utilisation de ces outils en routine dans les actions d'épidémio-surveillance faites à différentes échelles, dont celle du territoire, devrait nous permettre d'améliorer la précision des données sanitaires associées aux cultures de céréales.
Cependant, compte tenu du nombre croissant de travaux conduits sur ce pathosystème, il est probable que d'autres espèces, souches et/ou variants viraux impliqués dans la maladie du « pied chétif » soient prochainement identifiés dans les réservoirs sauvages et/ou les parcelles cultivées.
Leur incidence respective sur les cultures devra donc être établie.
Pour cela, il faudra de nouveaux outils et/ou techniques de diagnostic, adaptés aux caractéristiques de ces nouvelles entités virales.
Perspectives de recherche
Identifier les espèces de cicadelles et les souches de virus
Les travaux conduits par la communauté scientifique sur le pathosystème du « pied chétif » ont permis de mettre en évidence la complexité de ce modèle.
En effet, les données acquises révèlent qu'une situation d'apparence simple correspondant à la présence d'un symptôme de « pied chétif » sur céréale s'avère pouvoir dériver de différentes situations sanitaires en fonction de l'isolat de WDV considéré (souche/variant) et des espèces de cicadelles Psammotettix présentes sur la parcelle.
L'identification fine de ces deux partenaires (virus et vecteur) est donc indispensable à la gestion des risques sanitaires associés à la présence de « pied chétif » sur céréales. En effet, l'extension d'un foyer infectieux ayant émergé au sein de la parcelle dépend en grande partie :
– des caractéristiques intrinsèques de l'hôte, de la cicadelle et du virus,
– des interactions mises en place entre eux.
Prévoir l'extension des foyers, un enjeu pour l'avenir
À partir des connaissances actuelles (cf. ciavant), il est actuellement impossible d'anticiper la dynamique de l'extension des foyers infectieux à l'échelle de la parcelle.
Autrement dit, l'observation de symptômes et/ou de cicadelles Psammotettix dans un environnement céréalier est certes un paramètre incontournable pour appréhender le risque WDV à l'échelle de la parcelle, mais elle ne suffit pas pour appréhender le niveau du risque épidémique réel auquel est soumise la parcelle.
En effet, des situations sanitaires contrastées ont été décrites dans des environnements de culture similaires.
La présence de cicadelles Psammotettix, même virulifères, sur une parcelle ne conduit pas systématiquement à une situation sanitaire critique.
En fait, toute une série de facteurs vont conditionner l'installation du virus et moduler l'efficacité avec laquelle ce bioagresseur va se propager en colonisant, de plante en plante, l'ensemble de la culture.
Or, comme déjà écrit ci-avant, les données disponibles sur :
– la diversité de l'espèce WDV,
– la gamme des espèces cicadelles Psammotettix vectrices,
– la dynamique de la transmission virale et
– la variation intra-spécifique de la capacité vectrice des cicadelles...
... sont encore très partielles. Aussi, les enjeux des programmes de recherche en cours et à venir, sont précisément d'améliorer nos connaissances sur ces paramètres biologiques.
À terme, ces recherches ambitionnent de pouvoir disposer d'arguments permettant d'orienter et raisonner la stratégie de lutte contre ces phytopathogènes viraux responsables de la maladie du pied chétif, sur la base de la nature des espèces (vectrices/non vectrices) présentes, de la fréquence de cicadelles virulifères et de l'identité des souches virales diagnostiquées.
Fig. 1 : Localisation et intensité des foyers infectieux du WDV sur parcelles de céréales (données issues d'observations et d'analyses sérologiques effectuées au cours de la campagne 2007-2008).
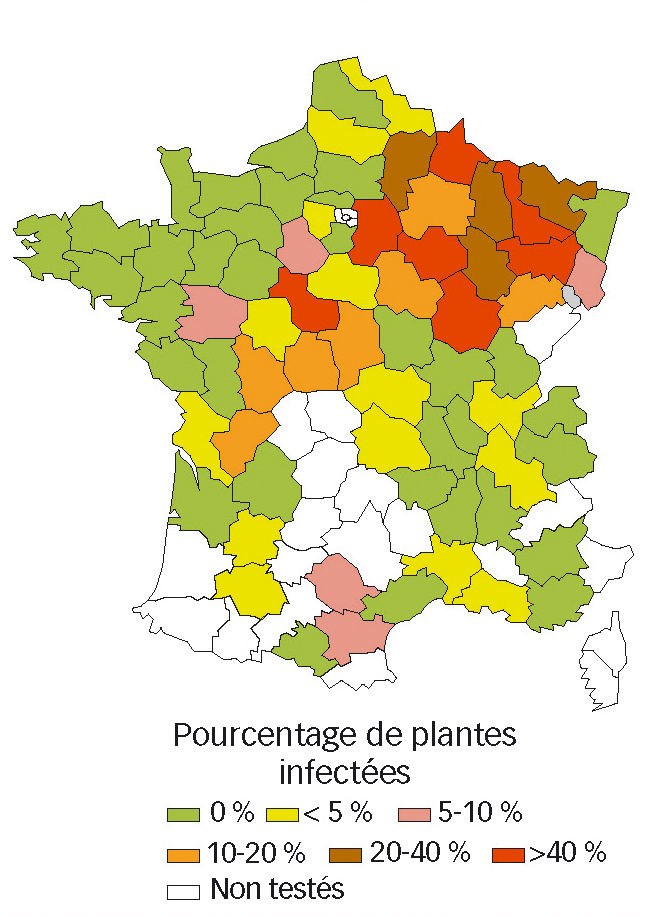
Fig. 2 : Représentation schématique des cycles biologiques d'une céréale d'hiver et de la cicadelle Psammotettix.
Les principales étapes du développement (du semis à la récolte) de la céréale sont présentées en vert. Les pointillés gris représentent la colonisation des graminées sauvages par les cicadelles après récolte. Les flèches brunes représententles cycles biologiques successifs des cicadelles. La durée de ces cycles (de l'œuf à la mort de l'adulte) est d'environ 71 jours (Guglielminoet Virla,1997). Les œufs pondus àla fin de l'automne éclosent à la sortie de l'hiver au moment de l'augmentation des températures et de la longueur des jours. La durée moyenne (à 20 °C, 70-95 % RH, 18 heures de lumière par jour) des 5 stades larvaires (L1 à L5) nécessaires à la formation des adultes est respectivement de 5.9, 5.1, 5.6, 6.3 et 9.4 jours (Manurung et al., 2005). La reproduction sexuée conduisant à la production d'œufs débute au 7e jour du stade adulte.
Fig. 3 : Classification et principales caractéristiques des groupes et variants du Wheat Dwarf Virus.
Les virus de la famille Geminiviridae sont répartis en quatre genres, Mastrevirus (membre type : Maize streak virus, MSV), Curtovirus (membre type : Beet curly top virus, BCTV), Begomovirus (membre type : Bean golden mosaic virus, BGMV) et Topocuvirus (membre type : Tomato pseudo-curly top virus, TPCTV). L'espèce Wheat dwarf virus (WDV) appartient au genre Mastrevirus. Les différentes souches de l'espèce WDV (WDV-bar[Tu], WDVa, WDVb et WDV-bar) partagent au minimum 75 % d'identité nucléotidique. La classification (famille, genre, espèce et souche) et les caractéristiques de ces différents niveaux de classification sont présentées.
* ODV : Oat dwarf virus, espèce décrite en 2007.
Fig. 4 : WDV, organisation de son génome.
Le cercle noir représente le génome du WDV Enkoping 1. Les gènes schématisés par des flèches sur le génome codent pour la protéine de mouvement (MP), la protéine de capside (CP) et les polymérases (REP et REP A). Ce génome présente également des régions intergéniques (LIR et SIR) impliquées dans la réplication et l'expression des protéines virales. Les positions nucléotidiques de ces régions sont indiquées sur le génome.
* Numéro d'accession GenBank : AJ311031.

