Imprimer
ImprimerLe puceron vert du pêcher Myzus persicae ne ravage pas seulement les pêchers. C'est en fait une espèce extrêmement polyphage infestant des cultures très variées, en particulier le colza. La nuisibilité de ce puceron découle de dégâts directs liés à sa prise de nourriture mais surtout à sa capacité à transmettre des viroses. Sur colza, il est le vecteur de trois virus plus ou moins dommageables (TuYV : virus de la jaunisse du navet, TuMV : virus de la mosaïque du navet, CaMV : virus de la mosaïque du chou-fleur). Jusqu'ici, malgré la présence d'une résistance aux insecticides de la famille des pyréthrinoïdes, il était maîtrisable grâce à d'autres familles d'insecticides. Mais la situation se complique...
Ce qu'on savait déjà au champ sur Myzus persicae et le colza
La situation dans le colza
Myzus persicae se caractérise par un cycle biologique complexe : il peut se reproduire de façon asexuée par parthénogenèse, ces phases asexuées alternant avec de rares cycles sexués, non obligatoires. Dans les populations se développant sur colza à l'automne, la reproduction asexuée est majoritaire.
Pour lutter contre ce puceron en culture de colza, les agriculteurs disposent aujourd'hui de trois familles d'insecticides : les carbamates, les pyréthrinoïdes et, depuis 2009, les néonicotinoïdes.
Pourquoi la lutte est-elle délicate ?
Cette lutte est délicate à gérer car les phénomènes de résistances aux insecticides chez M. persicae sont répandus. De plus, l'extrême polyphagie ainsi que le cycle de reproduction de M. persicae complexifient la dispersion des allèles de résistance dans les populations.
En effet, les pucerons peuvent arriver sur la culture en ayant déjà acquis un spectre de résistances à divers insecticides utilisés sur d'autres cultures hôtes et auxquels ils ont été exposés. Par ailleurs, le cycle biologique associant reproduction sexuée et asexuée permet à la fois des recombinaisons génétiques et une dissémination rapide des combinaisons les plus avantageuses, via la reproduction clonale.
Des résistances aux pyréthrinoïdes sur colza depuis 1997
Dans des populations de M. persicae collectées sur colza, les premiers cas de résistances aux pyréthrinoïdes ont été détectés à partir de 1997 en France (Source Cetiom-« Colza pucerons verts du pêcher et résistance aux pyréthrinoïdes »-Rencontres techniques régionales 2006-2007).
Pourtant jusqu'en 2005, la pullulation des pucerons verts sur colza est restée discrète grâce notamment à l'utilisation d'insecticides à base de pyrimicarbe (famille des carbamates).
Recrudescence depuis 2005, soupçon de nouvelles résistances
Mais depuis 2005, ce ravageur est de nouveau plus présent (Source Cetiom – « Colza pucerons verts du pêcher et résistance aux pyréthrinoïdes », Rencontres techniques régionales 2006-2007). Cela laisse suspecter une forte baisse de sensibilité des pucerons vis-à-vis des substances actives appliquées (pyréthrinoïdes et carbamates).
Ce qu'on savait déjà sur les mécanismes de résistance
Métabolique et « de cible », les deux grands types de mécanismes
Les résistances aux insecticides chez le puceron vert du pêcher (M. persicae) reposent sur deux grands types de mécanismes.
Des résistances de type métabolique. Souvent induites par la duplication d'un gène, elles engendrent une synthèse accrue de la protéine correspondante. La protéine surexprimée est une enzyme capable de dégrader une ou des substance(s) active(s).
Chez M. persicae, deux types d'enzymes sont connues pour être impliquées dans des résistances métaboliques : des carboxylestérases (E4 ou FE4) qui confèrent une résistance modérée vis-à-vis d'un large spectre d'insecticides (carbamates, pyréthrinoïdes, organophosphorés) (Field et al., 1993) et un cytochrome à P450 (Puinean et al., 2010) impliqué uniquement dans une résistance modérée aux néonicotinoïdes.
Des résistances dites de cible.
Elles sont dues à une modification de la protéine ciblée par l'insecticide et elles sont responsables d'une très forte baisse d'efficacité des insecticides concernés.
Trois grandes familles d'insecticides sont impactées par ce type de résistance : les pyréthrinoïdes, les carbamates et, plus récemment, les néonicotinoïdes.
Résistances de cible, situations variées par famille d'insecticides
Pour les néonicotinoïdes,une mutation responsable d'un haut niveau de résistance a été récemment décrite dans des populations vivant sur pêcher (Bass et al., 2011). Cette mutation affecte le gène codant pour le récepteur nicotinique, protéine cible de l'insecticide.
Concernant les carbamates, une mutation, responsable d'un fort niveau de résistance, a été identifiée sur le gène codant pour l'acétylcholinestérase 2 (Nabeshima et al., 2003), cible de cette famille d'insecticide. Cette mutation, nommée Mace pour « modified acétylcholinestérase », se traduit au niveau protéique par la substitution d'une serine par une phénylalanine au niveau de l'acide aminé 431 de l'acétylcholinestérase 2 (S431F).
Face aux pyréthrinoïdes, trois mutations peuvent être en cause chez M. persicae.
Deux de ces mutations sont bien connues. Elles affectent la cible de l'insecticide : le canal sodium. Elles diminuent l'affinité de l'insecticide vis-à-vis de sa cible. Ces résistances sont nommées kdr (knock-down resistance) et s-kdr (super kdr) (Martines-Torres et al., 1999).
La mutation kdr se traduit au niveau protéique par le remplacement d'une leucine par une phénylalanine en position 1014 de la protéine (L1014F). La mutation s-kdr se traduit par la substitution d'une thréonine à la place d'une méthionine en position 918 de la protéine (M918T) ; elle est toujours trouvée en association avec la mutation kdr (L1014F).
Une troisième mutation a été récemment décrite au niveau de ce codon correspondant à l'acide aminé 918 du canal sodium (Fontaine et al., 2011). Cette substitution se traduit au niveau de la protéine par le remplacement d'une méthionine par une leucine en position 918 du canal sodium (M918L). Cette mutation M918L, appelée ici s-kdr atypique, avait déjà été détectée chez le puceron du coton (Aphis gossypii).
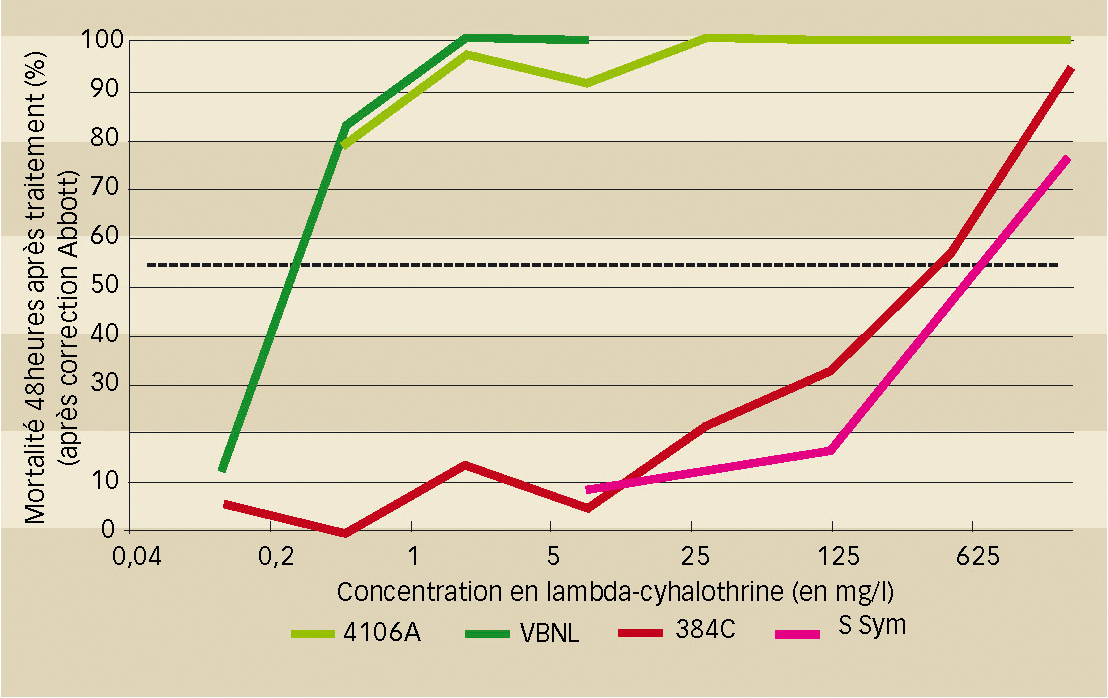
Les tests enzymatiques pour rechercher une résistance liée à des estérases et des tests biologiques réalisés au laboratoire par pulvérisation de pyréthrinoïdes (lambda-cyhalothrine) à l'aide d'une tour de Potter-Burgerjon ont clairement montré l'impact de cette mutation s-kdr atypique sur la perte de sensibilité des pucerons (figure 1 et Fontaine et al., 2011).
Notre travail pour évaluer la prévalence des résistances de cible sur colza
39 parcelles du quart nord-est
Nous avons cherché à évaluer la prévalence des résistances de cible aux pyréthrinoïdes et aux carbamates au sein des populations de M. persicae dans les cultures de colza du nord-est de la France.
Notre étude a porté sur environ 1 000 pucerons (80 % d'entre eux ont été échantillonnés en 2009 et le reste en 2010). Ils provenaient de 24 parcelles de production de colza et de 15 parcelles d'expérimentation.
Recherche par PCR
Pour la présentation du monitoring et la cartographie des mutations (Mace, kdr et s-kdr atypique M918L), lorsque plusieurs prélèvements de pucerons ont été réalisés sur le même site (cas des essais), seule la modalité TNT (témoin non traité insecticide) a été retenue. Les trois allèles de résistance (Mace, kdr et s-kdr atypique) ont été recherchés à l'aide de la technique PCR (Polymerase Chain Reaction).
Résistances de cible aux pyréthrinoïdes
Allèle kdr L1014F
L'allèle muté a été détecté dans 60 % des populations testées. Au sein des échantillons prélevés dans les parcelles de production, la proportion d'individus porteurs de l'allèle muté kdr est très variable (de 2,56 % à 80 % des individus analysés). Pour cinq parcelles, cette proportion est supérieure à 50 %. La plus forte occurrence de cet allèle de résistance (80 %) a été observée dans un prélèvement réalisé en Bourgogne sur culture non traitée. C'est en Ile-de-France (trois cultures sur quatre) et en Champagne-Ardenne (trois sur trois) que l'allèle kdr a été le plus souvent observé.
Pour les modalités non traitées des essais, la mutation a été détectée dans huit cas sur les douze. Il y a également une grande variabilité selon les essais (6 à 65 % des pucerons testés sont porteurs de l'allèle kdr).
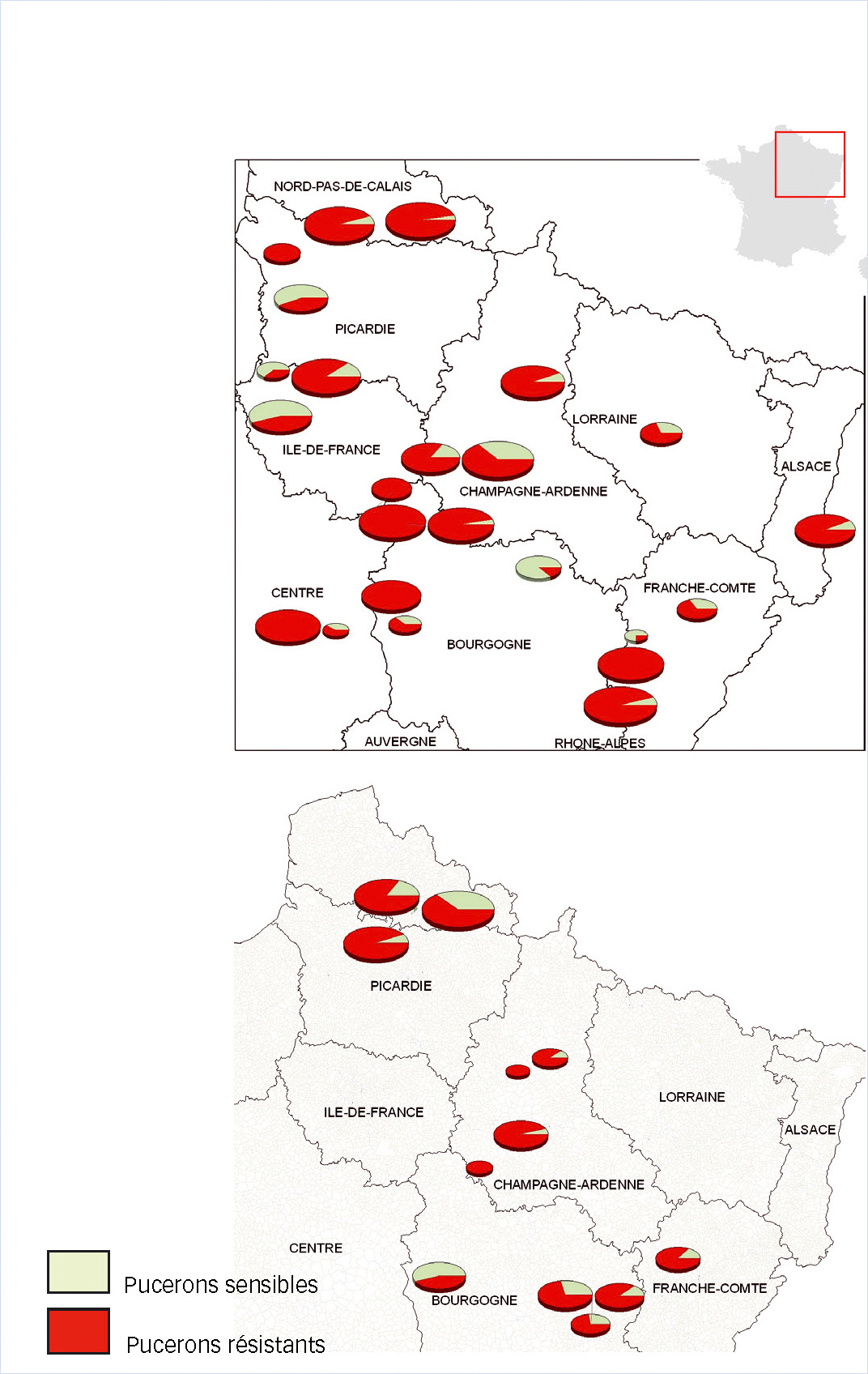
Allèle s-kdr atypique M918L (Figure 2)
L'allèle muté est présent dans toutes les parcelles/modalités échantillonnées. Selon les échantillons, il est détecté chez 13 à 100 % des individus testés. Pour 19 des 24 parcelles de production, il est rencontré chez au moins 50 % des individus testés. Toutes les régions sont concernées de façon semblable. Pour six parcelles, tous les individus testés sont porteurs de cet allèle (trois en Bourgogne et une pour chacune des régions suivantes : Centre, Franche-Comté et Picardie).
Sur les modalités d'essai non traitées, la situation est comparable à celle en parcelles de production : dans 11 cas sur 12, plus de la moitié des individus sont porteurs de l'allèle s-kdr atypique.
Résistance de cible aux carbamates (Mace)
La répartition de l'allèle muté Mace, responsable de la résistance aux carbamates, est identique à celle que nous avons observée pour s-kdr atypique M918L. Ceci s'explique par le fait que la quasi-totalité des pucerons porteurs de l'allèle muté s-kdr M918L sont également porteurs de l'allèle muté Mace.
Un génotype omniprésent !
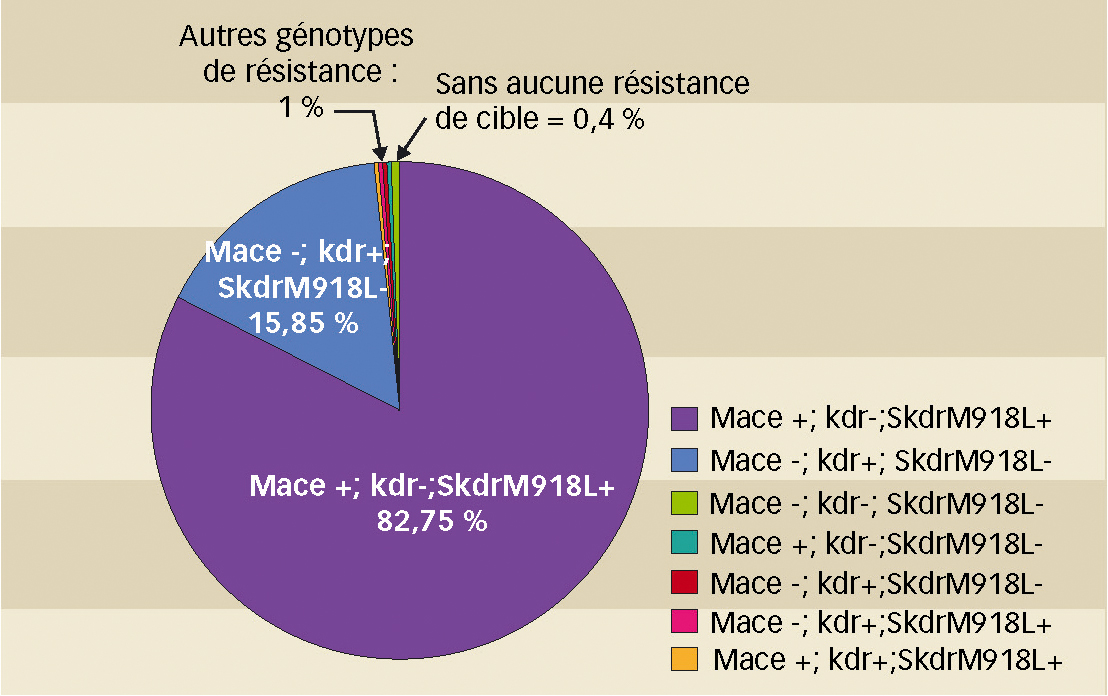
Résistances aux deux familles par « Mace + ; s-kdr atypique + » : 82,7 % des pucerons, partout
L'étude de l'association des allèles de résistance au sein d'un même individu révèle qu'environ 82,7 % des pucerons analysés pour les trois allèles (998 pucerons M. persicae) possèdent conjointement la mutation s-kdr atypique (M918L) et la mutation Mace, sans la mutation kdr (noté +/SS/RS sur la figure 3). La nouvelle mutation s-kdr atypique (M918L), toujours trouvée à l'état hétérozygote dans notre étude, se différencie bien de la mutation s-kdr classique (M918T) : elle n'est qu'exceptionnellement associée à la mutation kdr (seulement 0,2 % des individus testés possèdent les deux allèles kdr et s-kdr atypique).
Les autres génotypes
Concernant la mutation kdr, 15,9 % des pucerons (sur l'ensemble des régions) possèdent uniquement cet allèle de résistance (noté -/RS/SS sur figure 3). Comme pour l'allèle s-kdr atypique M918L, ils sont tous hétérozygotes pour cet allèle. Les deux génotypes cités ci-dessus (+/SS/RS et -/RS/SS) représentent donc plus de 98 % de pucerons analysés.
Les autres génotypes de résistance (1 % des pucerons analysés) se répartissent de la manière suivante : 0,1 % et 0,3 % des pucerons possèdent respectivement l'allèle de résistance Mace seul ou l'allèle s-kdr atypique seul ; 0,4 % possèdent les allèles Mace et kdr. Enfin, 0,2 % cumulent les trois allèles de résistance.
Une évolution prouvée
Comparaison entre 2009-2010 et 2001
L'ensemble de ces résultats permet de rendre compte de la présence et de la composition des allèles de résistance de cible aux carbamates et pyréthrinoïdes dans les populations de M. persicae infestant les cultures de colza.
Notre travail est à mettre en lien avec une précédente étude réalisée à partir de pucerons collectés à l'automne 2001 (Zamoum et al., 2005) sur des cultures de colza situées approximativement dans les mêmes régions. La comparaison des deux études montre que les allèles de résistance présents dans les populations de M. persicae ont fortement évolué en moins de dix ans.
Du génotype kdr, on est passé au « Mace + ; s-kdr atypique +»
Ainsi, un génotype résistant combinant deux mutations de cible associées à de très hauts niveaux de résistance aux carbamates et aux pyréthrinoïdes (respectivement Mace : S431F sur l'acétylcholinestérase 2 et s-kdr atypique M918L sur le canal sodium dépendant du voltage) s'avère omniprésent en 2009-2010 alors qu'en 2001, le génotype majeur était celui porteur uniquement de l'allèle kdr.
En parallèle, entre 2001 et 2009, l'allèle de résistance Mace est passé d'une fréquence inférieure à 1 % à une fréquence supérieure à 80 %.
Forte association entre allèles de résistance s-kdr atypique (M918L) et Mace
Raison de cette association, hypothèse « traitement »
La forte association entre deux allèles de résistance (Mace et s-kdr atypique) à deux familles d'insecticides différentes et le bouleversement de la fréquence des allèles de résistance dans les populations de M. persicae vivant sur colza sont surprenants et difficiles à expliquer sans études complémentaires.
Cette association entre deux allèles pourrait être la conséquence de pratiques phytosanitaires utilisant les deux familles chimiques en mélange pour la protection des cultures de colza.
Résistances dans les colzas non traités, hypothèse polyphagie
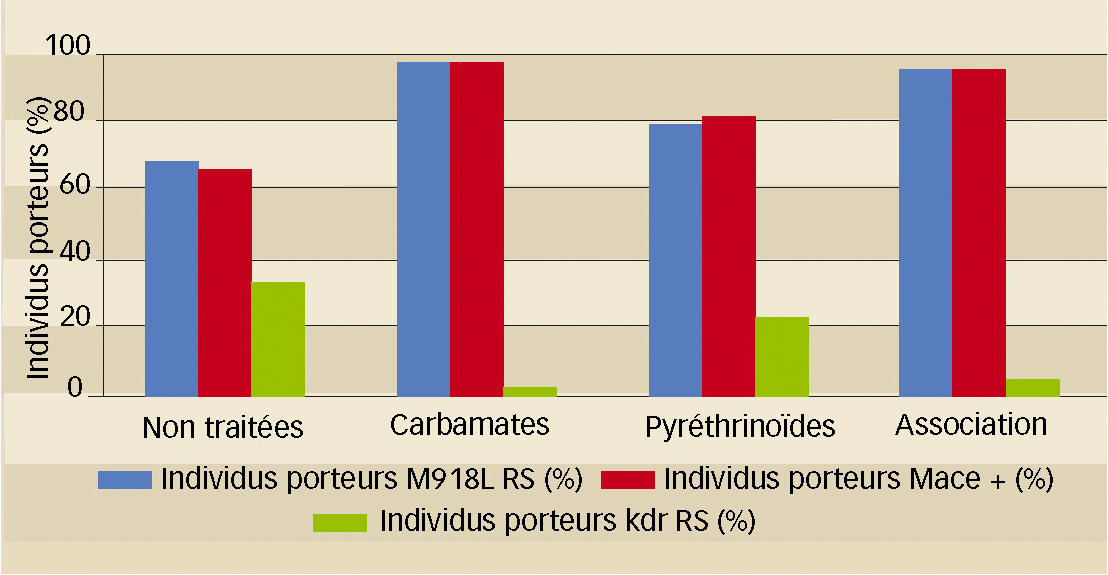
Le fait que plus de 80 % des individus testés dans notre étude soient porteurs des allèles Mace et s-kdr atypique indique également que la lutte avec les deux plus anciennes familles chimiques utilisées sur colza peut se révéler difficile.
En fait, ce génotype (Mace+ ; s-kdr atypique+) se retrouve à des taux élevés (68 %) même dans les parcelles témoins des essais (TNT) et les zones non traitées (ZNT) des parcelles commerciales (Figure 4), donc sans pression de sélection sur colza au cours de l'année de prélèvement.
L'extrême polyphagie de M. persicae pourrait expliquer en partie cette situation.
En effet, ce puceron est trouvé sur de nombreuses espèces d'arbres fruitiers et de plantes herbacées sauvages et cultivées. Comme les substances actives utilisées contre M. persicae, notamment sur betterave et pomme de terre, sont les mêmes que celles appliquées sur colza, les populations migrant sur colza pourraient avoir été soumises à des traitements insecticides avant d'arriver sur colza. Elles arriveraient avec leurs cortèges de mécanismes de résistance.
Le mode de reproduction des populations de M. persicae sur colza où les événements de recombinaison sexuée seraient rares et la parthénogenèse largement majoritaire – donc susceptible de multiplier à l'infini un génotype avantageux – pourrait également expliquer l'omniprésence du génotype Mace+ ; s-kdr atypique RS.
Conclusions-perspectives
Compter sur les néonicotinoïdes, tentant...
Ces résultats indiquent que les populations de M. persicae vivant sur colza sont très bien équipées en terme d'allèles de résistance vis-à-vis des deux anciennes familles d'insecticides autorisées sur cet usage : les carbamates et pyréthrinoïdes. La protection des cultures de colza repose désormais uniquement sur la nouvelle famille d'insecticides, les néonicotinoïdes, mise sur le marché en 2009 pour l'usage « Pucerons du colza ». Il est tentant de se contenter de ce mode d'action. Mais c'est imprudent.
... mais imprudent, vu la découverte récente d'une résistance de cible
En effet, le premier cas de résistance de cible vis-à-vis de cette famille chimique dans des populations de M. persicae prélevées dans des vergers de pêchers a été décrit en 2010 dans le sud de la France.
Cette famille est utilisée depuis 1994 sur pêcher pour lutter contre les pucerons. Des cas de résistance métabolique, sans grand effet sur l'efficacité au champ, avaient déjà été constatés. Mais la résistance de cible découverte en 2010 est responsable d'un niveau de résistance élevé (Bass et al., 2011) pouvant compromettre l'efficacité au champ des néonicotinoïdes. Certes les régions de culture du colza ne coïncident guère avec celles des vergers de pêchers. Mais les pucerons sont mobiles...
Au laboratoire, nos études vont donc se poursuivre sur les néonicotinoïdes afin de déterminer la prévalence et l'impact de la résistance de cible à cette famille chimique chez les populations de M. persicae vivant sur des plantes cultivées herbacées et ligneuses.
<p><b>Partenariat scientifique et technique :</b> Ce projet a été réalisé avec l'aide de Yannick Ballanger, du Cetiom, et grâce à la participation aux prélèvements du réseau des DRAAF-SRAL et du Cetiom.</p>