Imprimer
Imprimer
Femelle de Stenomesius japonicus en train de pondre sur une chenille de Tuta absoluta, elle-même visible en transparence dans sa mine. ph. A. Droui
, et les comptages aussi fiables grâce au confinement (filets). ph. A. Droui")
– Cages d'expérimentation sous serre utilisées pour le travail rapporté ici. Les conditions sont plus réalistes qu'au laboratoire (plantes-hôtes), et les comptages aussi fiables grâce au confinement (filets). ph. A. Droui
La protection contre Tuta absoluta, mineuse de la tomate, utilise la lutte biologique à l'aide d'auxiliaires. Mais aucun de ceux disponibles aujourd'hui n'agit contre les larves de T. absoluta. Nous présentons ici un essai sous serre d'un nouvel auxiliaire larvaire. Il montre la possibilité de l'utiliser en lutte biologique inoculative.
Pourquoi tester Stenomesius japonicus
Lutte contre Tuta, pourquoi biologique?
Face à l'arrivée de Tuta absoluta, il a fallu réagir rapidement. Rappelons que ce ravageur a envahi l'Europe en commençant par l'Espagne en 2006 (Desneux & al., 2011) et peut entraîner des pertes de 80 à 100 % dans les récoltes, si aucune mesure de lutte n'est entreprise (Desneux et al., 2010). Mais quel type de lutte ?
Actuellement, en serre, le contrôle des autres ravageurs de la tomate s'effectue de manière efficace grâce à la lutte biologique. L'utilisation d'insecticides contre T. absoluta risquerait de réduire l'efficacité des auxiliaires de culture. Par ailleurs le plan Ecophyto pousse à la recherche de méthodes alternatives, notamment l'usage d'auxiliaires. Il est donc urgent de mettre au point un programme de lutte biologique efficace contre ce nouveau ravageur.
Auxiliaires oophages déjà utilisés, mais pas les larvaires
Actuellement, le prédateur généraliste Macrolophus pygmaeus (Heteroptera : Miridae) et le parasitoïde oophage Trichogramma achaeae (Hymenoptera : Trichogrammatidae) sont déjà employés avec succès contre le stade oeuf de la mineuse (Chailleux et al., 2012). Mais on ne dispose pas encore d'auxiliaire permettant de maîtriser efficacement le stade larvaire, or ce sont les larves qui provoquent les dégâts.
Privilégier les auxiliaires indigènes, c'est conseillé et possible
L'étude des ennemis naturels indigènes doit être considérée comme prioritaire dans la lutte contre les ravageurs exotiques. En effet, l'introduction d'ennemis naturels exotiques n'est pas toujours vouée au succès. De plus, elle comprend des risques potentiels pour l'environnement et les autres espèces (Wajnberg et al., 2001).
Encore faut-il que des ennemis naturels indigènes existent. Autrement dit, que des espèces indigènes s'adaptent au nouvel hôte que représente un ravageur exotique.
C'est le cas pour T. absoluta. En effet, depuis son arrivée dans le bassin méditerranéen, un nombre croissant de prédateurs et parasitoïdes (parasites qui tuent leur hôte) indigènes attaquant ce ravageur a été observé. Cela suggère une adaptation rapide des ennemis naturels indigènes à l'invasion par T. absoluta (Desneux et al., 2010 ; Zappala et al., 2012).
Un parasitoïde trouvé sur le terrain, prometteur au laboratoire
Un ectoparasitoïde larvaire a été trouvé dans la zone paléarctique et notamment en France. Nommé Stenomesius japonicus (Hymenoptera : Eulophidae), il contrôle efficacement ce nouveau ravageur en laboratoire (voir encadré ci-contre).
Il était donc envisageable d'espérer l'utiliser contre T. absoluta en lutte biologique inoculative. Encore fallait-il le tester en conditions réalistes, c'est-à-dire sur plants de tomate mais aussi en présence d'un autre auxiliaire lâché par ailleurs dans les serres.
Pourquoi le tester combiné avec Macrolophus pygmaeus
Il y a des interactions entre auxiliaires dans les agro-écosystèmes
Bien que l'introduction d'une nouvelle espèce auxiliaire dans un système cultural soit souvent considérée comme améliorant l'efficacité globale de la lutte, son effet peut varier. La nouvelle espèce peut créer des interactions ou modifier les interactions entre ennemis naturels et ravageurs déjà présents. Il peut en résulter un contrôle plus efficace des ravageurs... mais parfois un contrôle plus faible !
Les interactions sont de deux types, directs ou indirects. Leur impact dépend de nombreux paramètres, notamment des caractéristiques biologiques des agents de lutte en jeu, mais aussi de l'échelle de temps considérée (Janssen et al., 1998).
En effet, si les mécanismes comportementaux prévalent à court terme, les mécanismes de dynamique des populations prennent le dessus sur le long terme, et leurs effets peuvent s'inverser (Rosenheim et Harmon, 2006).
Ces interactions peuvent conduire soit à la coexistence entre les différents agents de lutte, soit à l'exclusion de l'un d'entre eux (Janssen et al., 1998 ; Rosenheim et Harmon, 2006). La plupart des modèles théoriques prédisent que deux espèces en compétition pour une même ressource ne peuvent coexister qu'en présence de mécanismes réduisant le degré de compétition (Briggs, 1993).
Sur tomate, il faut prendre en compte le programme de lutte biologique préexistant contre les aleurodes
En France, en serre de tomates, l'agent de lutte dominant est la punaise prédatrice Macrolophus pygmaeus. Ce prédateur omnivore est utilisé en lutte biologique inoculative en particulier contre les aleurodes. Il consomme les oeufs et, à l'occasion, les jeunes larves de T. absoluta (Urbaneja et al., 2009). Il partage donc sa ressource avec le parasitoïde S. japonicus.
Cependant, les deux auxiliaires s'attaquant à des stades différents, la compétition pour la ressource s'effectue en amont. À cette compétition peuvent s'ajouter des interactions directes type prédation intraguilde si la punaise, très polyphage, consomme des larves de S. japonicus en train de parasiter des larves de T. absoluta.
Dans ce contexte, nous nous sommes attachés à déterminer si le parasitoïde larvaire était capable de subsister sous serre à long terme, soit seul, soit en présence du prédateur M. pygmaeus. Nous avons comparé son efficacité à celle du prédateur seul.
Une expérimentation en conditions réalistes
Trois combinaisons testées sur plants de tomate en cage sous serre
L'expérience s'est déroulée en serre verre sous conditions climatiques semi-contrôlées, en cage de 1 x 0,7 x 1 m (photo ci-dessus). Six plants de tomate ont été placés par cage, arrosés automatiquement avec une solution nutritive enrichie en NPK.
Trois combinaisons d'auxiliaires ont été testées sur deux niveaux d'infestation de T. absoluta (faible : 4 couples par cage ; élevé : 12 couples par cage). En fait, ce dernier paramètre s'est révélé sans impact sur la dynamique des populations. Aussi les deux densités testées seront donc regroupées dans la présentation des résultats. Les trois combinaisons sont les suivantes :
T. absoluta + parasitoïdes.
T. absoluta + parasitoïdes + punaises prédatrices.
T. absoluta + punaises prédatrices.
Lâchers et comptages
Les lâchers par espèce ont été réalisés en deux fois avec, à chaque lâcher, la moitié de la quantité totale d'individus, afin d'éviter les problèmes de populations artificiellement cycliques (Jaworski, 2011).
Le lâcher de M. pygmaeus (12 couples par cage au total) a débuté un mois avant les comptages afin de leur permettre de réaliser un cycle complet et de s'installer précocement dans la culture. Afin de favoriser l'installation des prédateurs, des oeufs d'Ephestia kuehniella ont été ajoutés dans les cages pendant deux semaines. Les premiers parasitoïdes ont été lâchés dès l'apparition de larves de T. absoluta de stade 2 à 3 (12 couples par cage au total), en deux lâchers (en début d'expérimentation), comme les autres espèces.
Les populations des trois espèces ont été échantillonnées à partir du 13 juin pendant 8 semaines. Pour chaque cage, 2 plantes ont été échantillonnées par semaine. Les plantes ont été observées alternativement chaque semaine, de manière à ce que chacune ait été échantillonnée. Les résultats ont été analysés statistiquement à l'aide du logiciel R (version 2.14.1).
Résultats
Coexistence possible sur la durée testée
Après 8 semaines de comptages, nous avons bien retrouvé des individus de S. japonicus et de M. pygmaeus dans les combinaisons avec les deux auxiliaires. Ceci prouve que la coexistence a été possible sur une période de plus de deux mois. Les populations ont eu une dynamique cyclique caractéristique des dynamiques proie-prédateur observées dans le milieu naturel.
Dynamique du parasitoïde larvaire, S. japonicus
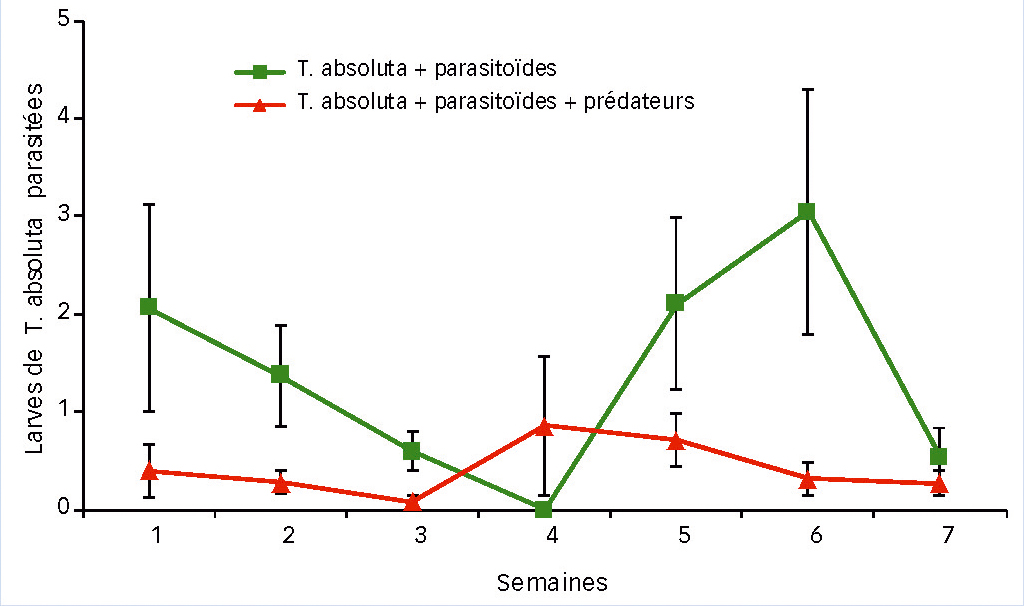
Dans la modalité parasitoïde seul, le parasitisme a baissé pendant la première moitié de l'expérience pour atteindre une valeur proche de zéro, puis il y a eu un pic important de trois larves parasitées par feuille en semaine 6 (Fig. 2). Lorsque les deux auxiliaires étaient présents (Fig. 2 également), le parasitisme par S. japonicus est resté bien plus faible (inférieur à 1 larve parasitée par feuille) et assez constant, malgré un léger pic en semaine 4. Les différences sont significatives (P < 0,001).
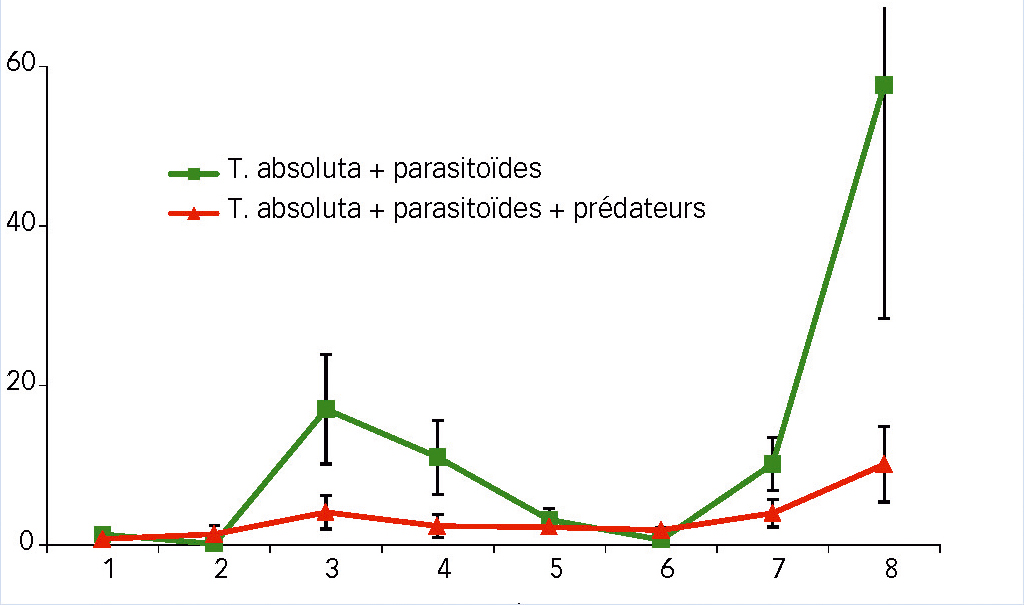
La dynamique des adultes de S. japonicus (Fig. 3) présente 2 pics dans la modalité parasitoïde seul, celui en fin d'expérimentation (semaine 8) montrant un nombre élevé d'individus par cage (57 en moyenne). La présence du prédateur M. pygmaeus a eu un effet négatif significatif sur la population d'adultes de S. japonicus (P < 0,001).
Dynamique de la punaise prédatrice, M. pygmaeus
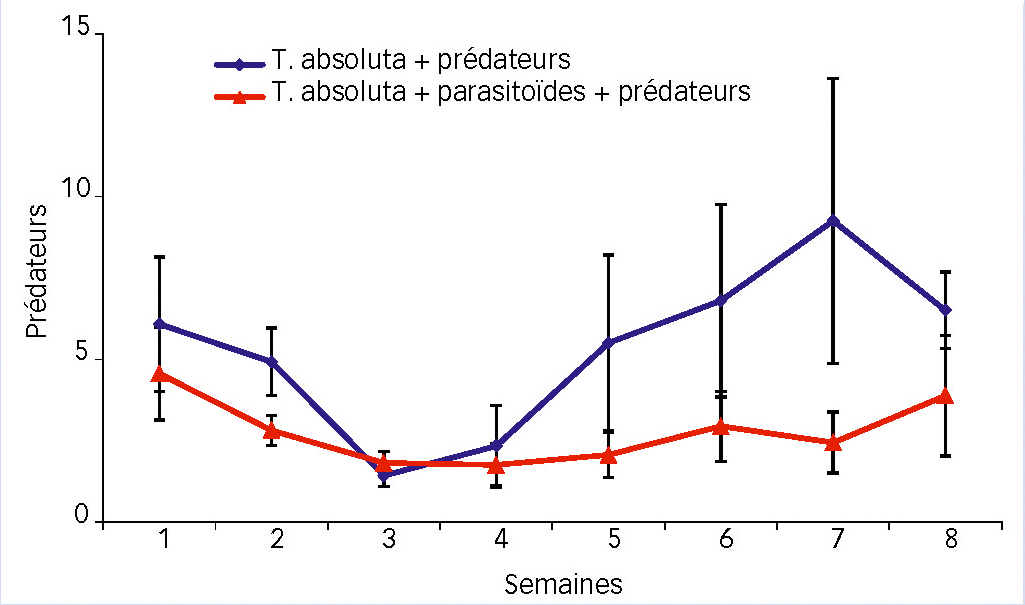
La dynamique de M. pygmaeus (Fig. 4) semble suivre la même évolution pour les deux modalités, avec une baisse pendant les trois premières semaines (fin d'un premier pic) puis une croissance de la population par la suite. Pour la modalité prédateur seul, le pic (en semaine 7) est bien plus prononcé qu'en présence de S. japonicus.
La présence du parasitoïde influence négativement la dynamique des populations du prédateur (P < 0,05). Cela se traduit par une certaine stagnation du nombre de M. pygmaeus par plante.
Ceci dit, pour les deux modalités, le nombre le plus faible de M. pygmaeus par plante (en semaine 3) ne chute guère en dessous des deux individus par plante qui est la dose initiale de lâcher recommandée.
Ravageur, T. absoluta, point commun aux trois modalités
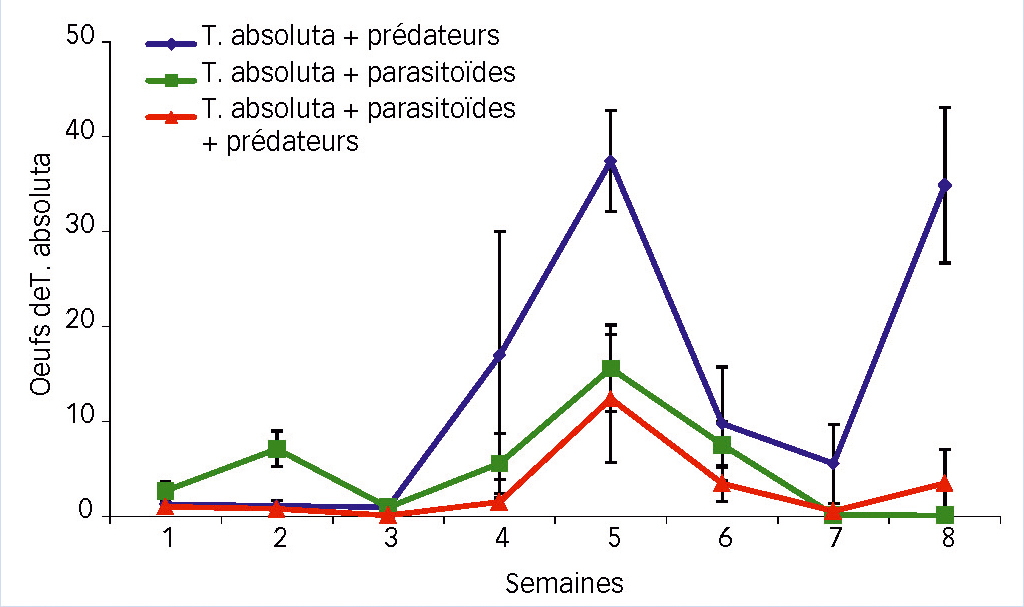
Sur la période de comptage, on observe un premier pic d'oeufs en semaine 2, un second plus important en semaine 5 et l'esquisse d'un troisième en semaine 8 (Fig. 5).
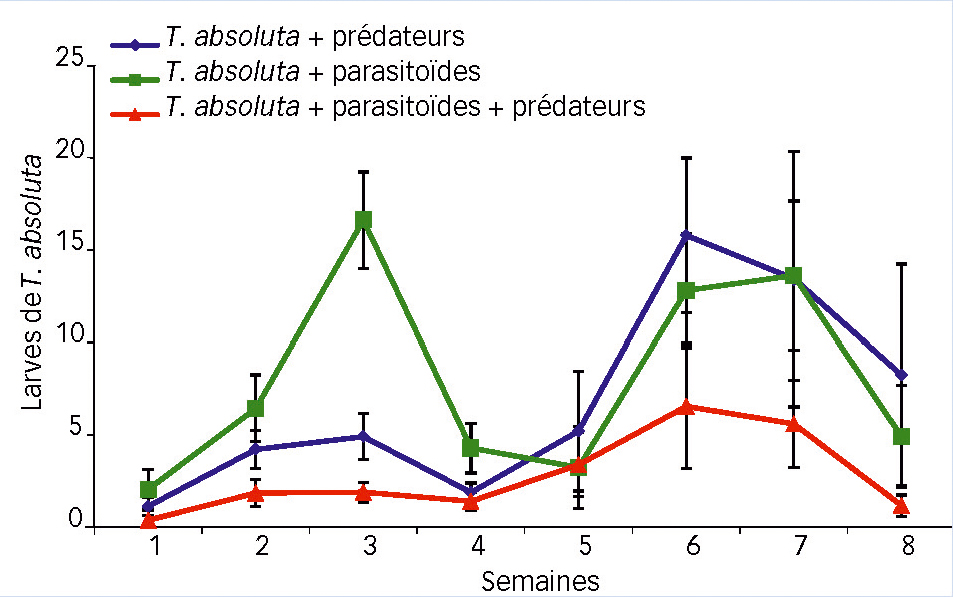
La dynamique des populations de larves de T. absoluta présente deux pics pendant la période, atteints en semaine 3 puis en semaines 6 à 7 (Fig. 6).
Face à un auxiliaire seul
La comparaison de la dynamique des oeufs de T. absoluta (Fig. 5) montre des valeurs extrêmes de 38 oeufs de ce ravageur par feuille pour la modalité M. pygmaeus seul, plus de deux fois supérieures à celles des deux autres modalités. L'écart se creuse encore davantage avec le troisième pic de population (semaine 8). Ceci suggère un contrôle insuffisant par le prédateur M. pygmaeus seul.
Par ailleurs, dans la modalité avec S. japonicus seul, le nombre de larves est significativement plus élevé lors du premier pic (semaine 3) que dans la modalité avec les deux auxiliaires combinés (Fig. 6).
Face à la combinaison des deux auxiliaires
On le voit sur la figure 5, l'association des deux auxiliaires dans la modalité les combinant a un impact très significatif sur la quantité d'oeufs du ravageur (P < 0,001).
Sur l'ensemble de l'expérimentation, de même que pour les oeufs de T. absoluta, le contrôle des larves est plus efficace en présence des deux auxiliaires. C'est particulièrement visible pendant les trois dernières semaines (Fig. 6).
Pendant la deuxième moitié de la période d'observation, la densité de larves ne diffère pas entre les modalités avec le prédateur seul et le parasitoïde seul. Les auxiliaires présents contribuent de manière significative à expliquer les différences de populations observées chez les larves du ravageur (P < 0,01).
Dégâts sur la production
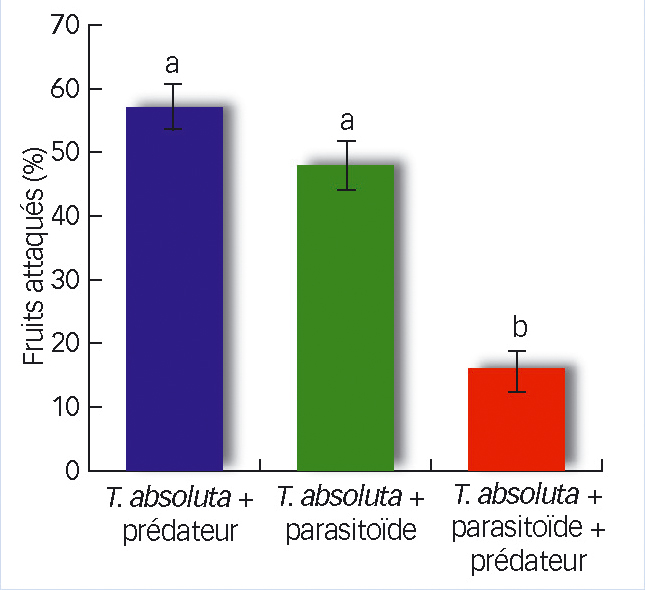
Le pourcentage de fruits attaqués par T. absoluta était significativement plus faible (p<0,05, et même dans ce cas p<0,01) quand les deux auxiliaires étaient ensemble. Ce résultat est la conséquence logique du meilleur contrôle des larves de T. absoluta en présence des deux auxiliaires observés précédemment (Fig. 7).
Des résultats encourageants
Combinaison : meilleur contrôle malgré la compétition
Nous avons mis en évidence un effet négatif réciproque entre les deux agents de lutte M. pygmaeus et S. japonicus (compétition pour la ressource). Pour autant, l'expérience a démontré que la coexistence était possible sur le long terme (plus de 3 mois d'expérimentation, périodes de lâcher et de comptage comprises) et que la présence combinée des deux auxiliaires fournissait un meilleur contrôle que chacun isolément.
Comprendre la coexistence des deux auxiliaires
Si de nombreux auteurs ont étudié la coexistence entre espèces par une approche de modélisation, le nombre d'études expérimentales dans ce domaine reste encore limité (Amarasekare, 2003).
La nôtre démontre que deux espèces partageant une ressource commune peuvent coexister sur le long terme.
Par ailleurs, la prédation intraguilde, certes observée en laboratoire (il s'agit ici de la consommation de larves de T. absoluta parasitées par le prédateur, conduisant à la mort du parasitoïde juvénile) n'a pas non plus gommé l'effet positif de l'association des deux auxiliaires sur le contrôle du ravageur. Cette observation avait été faite précédemment sur d'autres types de cultures (voir par exemple Snyder et Ives, 2003 ; ou Messelink et al., 2012).
L'effet bénéfique d'une combinaison prédateur/ parasitoïde peut s'expliquer dans notre cas par l'existence d'une niche limitant la compétition entre les deux espèces (partitionnement de la ressource selon le stade consommé ou l'axe de la plante par exemple) ou d'un compromis entre les caractéristiques biologiques liées aux stratégies alimentaires, spécialiste et généraliste, comme souligné par Symondson et al. (2002).
Lorsque les deux auxiliaires sont utilisés simultanément, le contrôle immédiat par M. pygmaeus réduit la pression de T. absoluta et permet l'installation progressive des populations de parasitoïdes.
Efficacité à confirmer en présence d'aleurodes
Cependant, l'agro-écosystème considéré pendant cette étude était simplifié. En particulier, il existe d'autres ravageurs et prédateurs en serre de tomates, notamment deux espèces d'aleurodes. Il se peut donc que les interactions observées soient modifiées dans un système plus complexe.
Plus particulièrement, il a déjà été démontré que la présence de la proie alternative Trialeurodes vaporariorum fournissant du miellat, consommé par les parasitoïdes, impactait positivement leur longévité (Riggi, données non publiées, 2012). Cette expérimentation contribue à souligner la complexité des interactions dans le cadre de la lutte biologique et démontre les effets bénéfiques, dans le cas étudié, de l'association de plusieurs auxiliaires contre un ravageur.
Perspectives en terme de lutte biologique inoculative
Stenomesius japonicus pour renforcer l'action de M. pygmaeus
D'un point de vue pratique, on a bien retrouvé chez M. pygmaeus les caractéristiques d'un prédateur généraliste exerçant un contrôle efficace en début d'infestation, et chez S. japonicus les attributs d'un spécialiste fluctuant suivant les densités de proies et plus efficace sur la durée.
Si le prédateur est déjà utilisé de manière efficace contre T. absoluta, il n'a pas permis à lui seul de contrôler le ravageur dans nos expérimentations. En revanche, le contrôle a été plus efficace en combinant les deux agents de lutte, dont l'effet a été additif.
De plus, S. japonicus ayant persisté longtemps après un lâcher inoculatif, il est donc envisageable de l'utiliser en lutte par inoculation contre T. absoluta, en combinaison avec M. pygmaeus.
S. japonicus est un parasitoïde indigène adapté à notre climat. Il constitue donc un bon complément à la lutte biologique avec M. pygmaeus, ce dernier étant incapable de maintenir à lui seul le ravageur sous des seuils économiques de nuisibilité.
Précautions à prendre et points restant à étudier
Mais, des dégâts non négligeables de T. absoluta ayant été tout de même constatés dans certaines répétitions, il est indispensable d'introduire le prédateur le plus tôt possible. Cela permettra de limiter au maximum les dégâts et maintenir les populations du ravageur à des densités basses plus facilement contrôlables. Le parasitoïde pourra ensuite être inoculé dès l'observation des premières larves en saison. Il reste encore à définir les doses de lâcher sur le terrain.
La mise en place d'une stratégie de lutte efficace et économiquement viable est indispensable afin de stopper la progression de ce ravageur invasif. Notre étude, basée entièrement sur de la lutte biologique, apporte des résultats encourageants dans cette direction.
<p><b>Remerciements :</b></p> <p>INRA, Départements « Santé des Plantes et Environnement » (2009-2010) et « Environnement & Agronomie » (2010-2012).</p> <p>InVivo AgroSolutions, Groupe InVivo.</p> <p>ANRT et Biotop (Groupe IAS) (Bourse CIFRE, 2010-2013).</p> <p>Ministère de l'Agriculture (Projet Casdar TutaPI, 2011-2013).</p>
Le parasitoïde larvaire, ce qu'on en a appris au laboratoire

Fig. 1 : Cycle biologique de S. japonicus.
Nous avons élevé Stenomesius japonicus au laboratoire. Les femelles peuvent y vivre jusqu'à 60 jours. Elles pondent en moyenne 200-250 oeufs au cours de leur vie (moyenne de 1,5 oeuf par larve de T. absoluta), avec une fécondité maximale une semaine après l'émergence. En moyenne et dans nos conditions de laboratoire, une femelle peut tuer 87 larves d'hôtes (pour se nourrir) et en parasiter 151 autres (pour la ponte) pendant sa vie. Le parasitisme est le plus élevé pendant les 30 premiers jours de vie de l'imago.
Par ailleurs, les larves de S. japonicus sont très mobiles et capables de passer d'un hôte à l'autre, au moins à l'intérieur d'une même mine. La figure 1 schématise le cycle de S. japonicus.