 Imprimer
Imprimer et de maïs (photo 3). Tous deux porteurs de périthèces de F. graminearum. Photos : E. Moreau")
1- Fusariose sur épi de blé tendre. 2 et 3 - Deux résidus de culture étudiés : débris de blé tendre (photo 2) et de maïs (photo 3). Tous deux porteurs de périthèces de F. graminearum. Photos : E. Moreau
La fusariose de l'épi, majoritairement causée par F. graminearum et Microdochium spp., est l'une des maladies les plus préjudiciables du blé. Elle occasionne des pertes de rendement, mais aussi une dépréciation de la qualité technologique et sanitaire des grains.
F. graminearum produit des mycotoxines dangereuses pour la santé humaine et animale, dont le déoxynivalénol (DON) et la zéaralénone (ZEA), réglementées depuis 2006 au niveau européen pour les denrées destinées à l'alimentation humaine.
Le respect des limites réglementaires étant nécessaire à la commercialisation du blé, mieux connaître l'agent producteur est utile pour la mise en place d'une lutte efficace.
Étudier la phase saprophyte du pathogène, pourquoi ?
Cycle à deux phases, ce qu'on savait déjà
Le cycle de développement du champignon pathogène F. graminearum a été très étudié ces dernières années (Trail, 2009). Il comprend deux phases : pathogène et saprophyte (Figure 1).
La première phase commence lors de l'infection de la plante au printemps et se traduit par la colonisation des tissus, le développement de symptômes sur la plante-hôte et la production de mycotoxines dans les grains. Le maïs, le blé dur et le blé tendre sont des plantes hôtes reconnues de F. graminearum (Pereyra & Dill-Macky, 2008).
La deuxième phase, moins bien connue, commence après la maturité des grains et se poursuit sur les résidus de culture restant au sol après la récolte. Elle constitue la forme de survie du champignon durant l'hiver.
Phase saprophyte, importance sûre... mais références contradictoires
Cette survie, essentiellement sous forme de périthèces (phase sexuée), serait la source majeure d'inoculum primaire susceptible d'infecter la culture suivante (Sutton, 1982). En France, des enquêtes réalisées depuis douze ans auprès d'agriculteurs, couplées à des études expérimentales, ont permis d'identifier et hiérarchiser les facteurs de risque de production de DON (Gourdain et al., 2009).
Le précédent cultural apparaît comme l'un des principaux, au vu de la capacité de survie saprophyte de F. graminearum sur les résidus de culture durant l'hiver.
La nature des précédents semble jouer un rôle notable dans le développement de la maladie car ils ne présentent pas tous le même risque. Mais... Les grilles de risque des divers pays européens présentent des disparités. Ainsi le précédent blé dur est classé à risque faible en France et à risque élevé en Italie.
Depuis 2010, des travaux engagés au sein d'Arvalis visent à mieux comprendre la phase saprophyte de F. graminearum, à travers l'étude du développement de ce champignon sur différents résidus de culture.
Au laboratoire et au champ
En conditions contrôlées (laboratoire), suivi de la phase sexuée de F. graminearum sur huit résidus
Le cycle de développement de F. graminearum étant très dépendant du climat, un essai en conditions contrôlées permet d'étudier plus finement la phase saprophyte du champignon.
Huit résidus de culture sont stérilisés puis inoculés avec F. graminearum et incubés vingt jours en conditions optimales au développement de sa reproduction sexuée, soit 20 °C, 80 % d'humidité relative et alternance 12 h jour/nuit. Il s'agit de résidus de betterave, blé dur, blé tendre, colza, maïs, pois, pomme de terre et tournesol. Les observations effectuées prennent en compte les structures reproductrices sexuées (périthèces) de F. graminearum.
L'objectif est triple :
– déterminer si la méthode d'analyse par PCR quantitative des résidus est réellement représentative de leur potentiel infectieux représenté par les périthèces,
– hiérarchiser ces résidus par le comptage des structures reproductrices,
Au champ, potentiel infectieux de deux résidus de culture
Pour compléter l'essai en conditions contrôlées, un essai a été réalisé en plein champ afin de tester le pouvoir infectieux de deux résidus de culture : blé tendre, maïs.
Ces derniers sont inoculés et placés au sol sous un blé tendre sensible à la fusariose (note GEVES : 3,5) et brumisé durant la période de floraison. Les résidus sont prélevés dans les micro-parcelles (quatre répétitions par précédent) à diverses dates pour une analyse de la quantité d'ADN fongique. À la récolte, des analyses d'ADN fongique et de mycotoxines sont réalisées sur grains.
L'objectif est de déterminer si les observations réalisées sur les résidus en conditions contrôlées sont transposables au champ, et de voir comment cela se traduit sur les grains de blé récoltés.
Résultats sur l'observation des résidus
Périthèces sur tous les résidus...
Lorsque les résidus sont inoculés et placés en conditions optimales pour initier la reproduction sexuée de F. graminearum, le champignon apparaît capable de se développer et produire des périthèces sur tous les résidus (Tableau 1).
Cependant, ce développement diffère selon le type de résidus « support ».
Après vingt jours d'incubation, on peut classer les résidus en deux groupes statistiquement homogènes selon le nombre de périthèces développés. Ainsi, le résidu de pomme de terre est très favorable au développement de périthèces, alors que celui de tournesol l'est bien moins.
Bien que le classement obtenu soit à relativiser au regard de la durée insuffisante de l'expérimentation, le développement différencié confirme des observations précédentes.
L'étude de la cinétique d'apparition des périthèces immatures présente différents profils selon les résidus (Figure 2). Ces résultats laissent supposer que F. graminearum se développe à une vitesse différente selon le type de résidus, vitesse qui peut être liée à la composition du résidu.
Pas de relation entre nombre de périthèces et quantité d'ADN
Le comptage des périthèces étant chronophage et exigeant une expertise, la question d'évaluer un potentiel infectieux de résidus par PCR-Q est de rigueur. Cette étude a été réalisée sur les résidus à la fin de l'essai.
Au final, en prenant des morceaux de résidus présentant une gamme de quantité de périthèces, aucune relation n'est observée entre les deux variables (Figure 3).
Cela semble relativement logique puisque l'ADN fongique comptabilise toutes les formes du champignon mortes ou vivantes : mycélium, périthèces et sporodochies (forme de reproduction asexuée).
Ainsi, la quantité d'ADN fongique d'un résidu ne semble pas être un bon indicateur de son potentiel infectieux réel. Néanmoins, elle reste un indicateur de la présence de F. graminearum ; c'est cet indicateur qui a été évalué sur les résidus épandus dans les parcelles de blé tendre en plein champ.
En plein champ, quantification de F. graminearum
Au champ, il apparaît que les résidus ne présentent que très peu d'ADN fongique lors de leur récolte (< 1pg/ng d'ADN total) (Figure 4). Cependant, si les valeurs stagnent pour le blé tendre tout au long de la saison, pour le maïs en revanche, elles présentent une importante variation.
L'augmentation est notable au prélèvement sortie hiver qui correspond à la fin du mois de mars et, à la floraison, les résidus de maïs présentent une quantité d'ADN de F. graminearum environ vingt fois plus important que les résidus de blé.
Que ce soit en conditions contrôlées ou au champ, F. graminearum semble ainsi présenter un développement différencié selon le type de résidus. Quels sont donc les facteurs qui pourraient expliquer ce différentiel ?
Impact des caractéristiques des résidus
Leur taille (élément de qualité) et leur quantité, réputées significatives
Un sondage réalisé en région Centre en 2010 s'était intéressé aux facteurs discriminant les résidus de culture entre eux, tels que leur taille ou leur taux de recouvrement.
Ainsi, il apparaissait que des résidus de taille supérieure à 5 cm semblaient plus favorables au développement de F. graminearum que ceux de taille plus réduite. Ces débris, moins vite dégradés, peuvent être un support possible pour F. graminearum.
La quantité de résidus restant au sol après récolte est importante. Cette quantité dépend du type de culture, du rendement et du type de travail du sol. Ainsi, pour un même itinéraire technique, des résidus de maïs et de colza ne seront pas présents en même quantité. Ceci pourrait expliquer les différences de risque entre les résidus.
Qualité des résidus, importance vérifiée
Pour répondre à ce questionnement, des mêmes quantités de résidus de blé tendre et de maïs ont été placés dans des micro-parcelles de blé tendre. On teste ainsi la qualité et non pas la quantité des résidus.
Les résultats montrent une différence de contamination des épis selon le type de résidus. Cela indique que la qualité des résidus est importante, de même que leur résistance à la dégradation. Au cours du temps, les résidus subissent en effet l'action de la microflore et microfaune du sol, qui vont modifier leurs caractéristiques physiques (diminution de taille) et chimiques (diminution de la teneur en azote).
Fusarium graminearum ne peut effectuer son cycle complet que si sa phase saprophyte s'effectue sur un résidu relativement résistant. Cette résistance à la dégradation va être liée à sa taille, sa composition biochimique (lignine, C/N…) ainsi qu'à sa teneur en eau.
La teneur en eau, nécessaire mais pas suffisante au laboratoire
Il est facile d'imaginer que la teneur en eau des résidus joue un rôle important dans le développement du champignon, qui a besoin d'une forte humidité pour survivre (Dufault et al., 2006). De précédents essais en champ semblaient indiquer l'importance de facteurs tels que l'humidité et la teneur en azote des résidus pour le développement du champignon. Les essais présentés ici tentent de préciser l'impact de ces facteurs.
En conditions contrôlées, il apparaît que la teneur en eau des débris est une condition nécessaire mais pas suffisante pour expliquer le développement de périthèces (Figure 5). En effet, les résidus très humides présentent deux cas de figures : certains en développent beaucoup (ex. betterave) et d'autres peu (ex. tournesol).
Teneur en azote, liée mais pas trop
La teneur en azote des résidus semble aussi jouer un rôle dans le développement du champignon, lequel a besoin d'une source azotée pour se nourrir (Trail et al., 2002). Une analyse en composante principale (ACP) montre que ce facteur évolue dans le même sens que les quantités de périthèces. Les variables « azote » et « périthèces » seraient donc liées sans pour autant présenter une corrélation significative (Figure 6).
Ainsi, F. graminearum est présent sur le résidu laissé au sol et, s'il est toujours présent à la floraison des blés (stade d'infection), il est intéressant de regarder si le taux de contamination des résidus reflète celui des épis.
Relation résidus/qualité du grain, les débris de maïs impliqués
Les analyses de PCR-Q sur résidus au champ indiquent des résidus de maïs plus contaminés que ceux de blé tendre lors de la floraison des blés. Certes, les résultats en conditions contrôlées indiquent que l'analyse de PCR-Q n'est pas un bon indicateur du potentiel infectieux (périthèces) : une plus forte quantité d'ADN de F. graminearum sur résidu de maïs n'entraîne pas forcément un inoculum plus important. Pourtant, l'analyse sur culture semble montrer un lien : intensité de maladie et impact sur le rendement plus importants en présence de résidus de maïs (Figures 7a et 7b).
Les analyses de grains valident le rôle majeur des résidus de maïs dans la contamination des grains par le champignon. Les quantités d'ADN de F. graminearum et de mycotoxines sont en effet majoritaires sur les blés à précédent maïs (Figures 7c et 7d). Ceci confirme que ce résidu est une source importante d'inoculum pour la culture suivante.
Conclusions et perspectives
En contamination forcée, F. graminearum est capable de se développer en saprophyte et de réaliser une reproduction sexuée par production de périthèces sur tout type de résidu, sans distinction entre plantes hôtes et non hôtes de sa phase pathogène.
Ce développement s'effectue à des degrés et des vitesses variables selon le résidu. Ce différentiel peut être mis en relation avec les caractéristiques des résidus, notamment teneur en eau et teneur en protéines.
Cependant, au champ, le développement saprophytique de F. graminearum est suivi non pas par le dénombrement des périthèces mais par des analyses de quantités d'ADN. L'étude réalisée ici montre que cette méthode ne permet pas de traduire le potentiel infectieux du résidu mais reste un indicateur de leur taux de contamination.
Enfin, ce taux de contamination semble relativement bien traduire la contamination réelle des épis par la maladie.
En effet, si on revient au schéma épidémiologique complet, il faut que diverses conditions soient réunies pour que la contamination des épis ait lieu. L'inoculum doit être présent sur les résidus du précédent, lesquels doivent être présents lors du stade sensible de la culture, et les conditions climatiques doivent être favorables à l'infection.
Pour éclairer la place de la phase saprophyte du champignon dans l'épidémiologie, il faudrait étudier la dégradation des résidus. Cela permettrait de pondérer le risque potentiel qui leur est lié (efficience du développement de F. graminearum).
Il serait intéressant, par ailleurs, de mieux déterminer la part de la reproduction sexuée et asexuée du champignon en phase saprophyte dans le développement de la maladie. Des recherches sont actuellement en cours sur ces deux sujets.
Fig. 1 : Cycle de Fusarium graminearum.
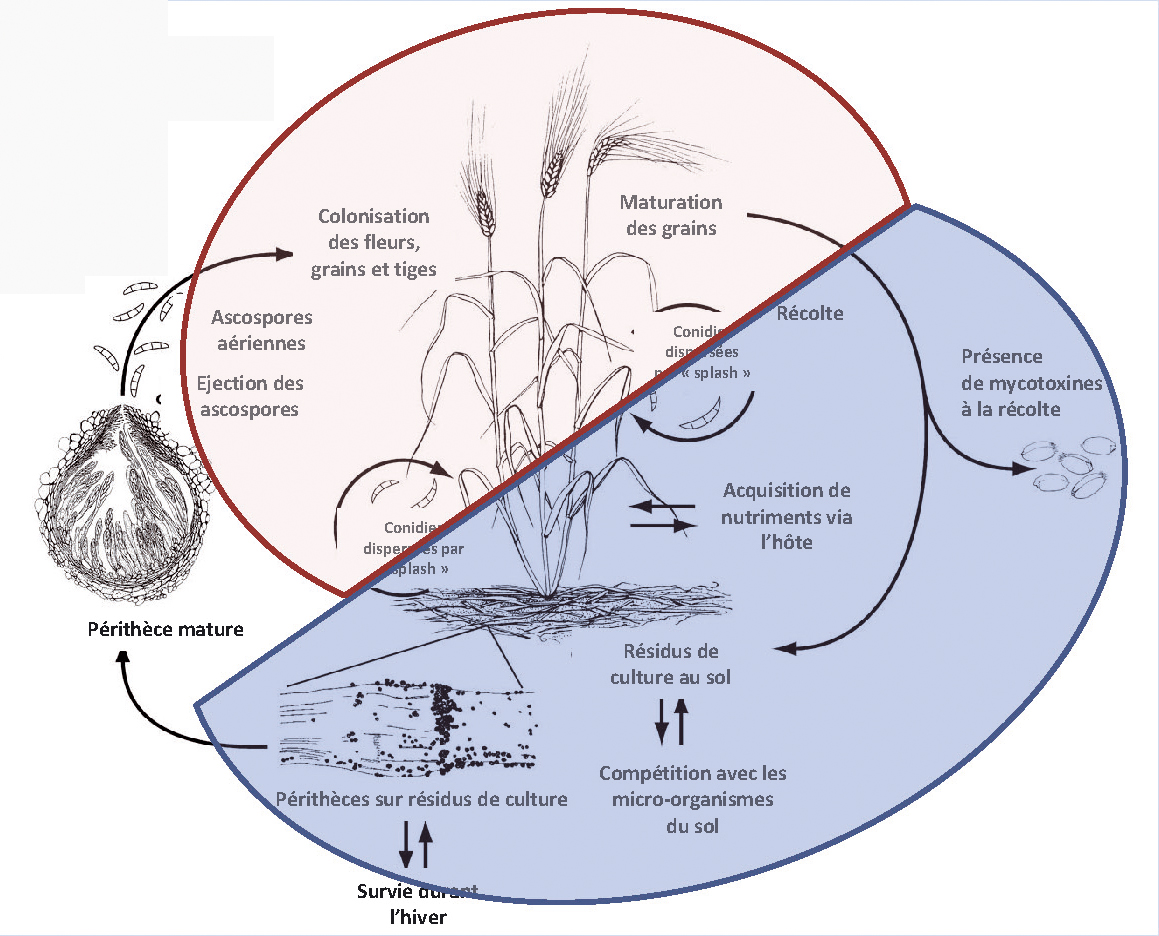
En bleu, la phase saprophyte où la ZEA est produite. En rose, la phase pathogène où le DON est sécrété. Source : Trail, 2009.
Tableau 1 : Développement des périthèces sur les différents résidus de culture.
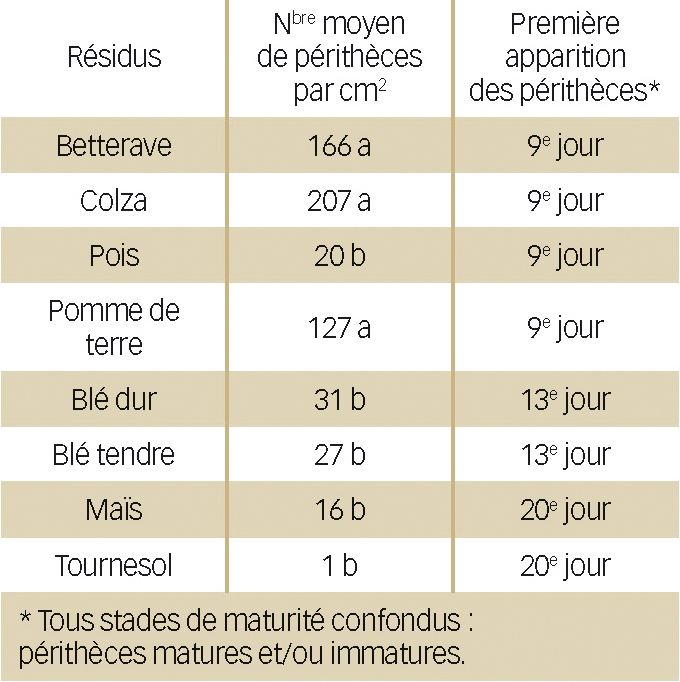
(Les lettres a et b représentent des groupes significativement différents, α = 5 %, test de Tuckey).
Fig. 2 : Au laboratoire, la nature du résidu
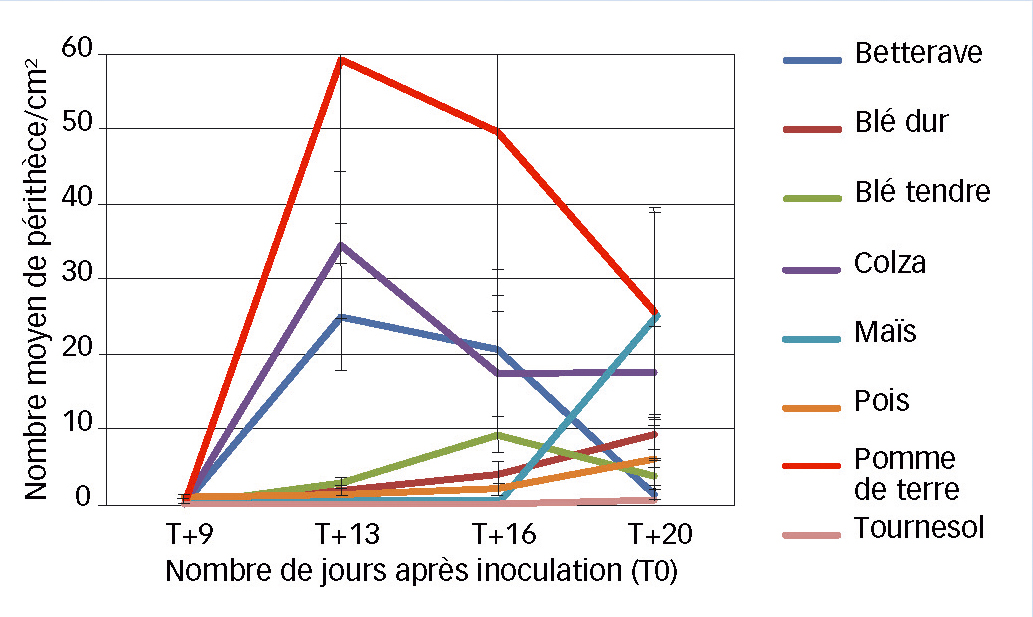
La cinétique d'apparition des périthèces immatures diffère selon l'espèce végétale du résidu « support ».
Fig. 3 : Au laboratoire, pas de lien entre ADN et périthèces
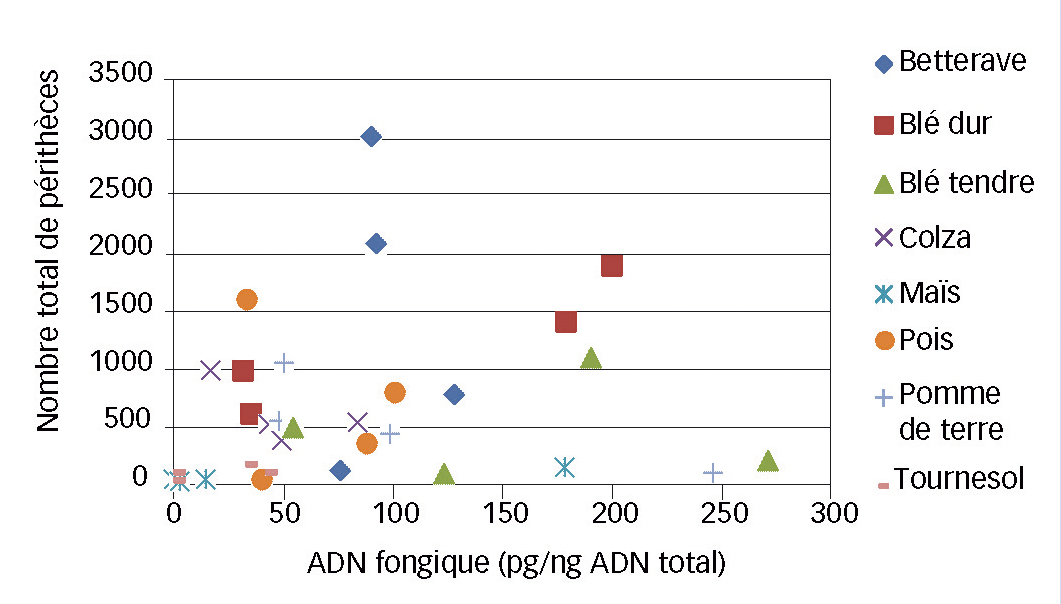
Relation entre le nombre de périthèces et la quantité d'ADN fongique.
Fig. 4 : Au champ, les résidus de maïs n'évoluent pas comme ceux de blé
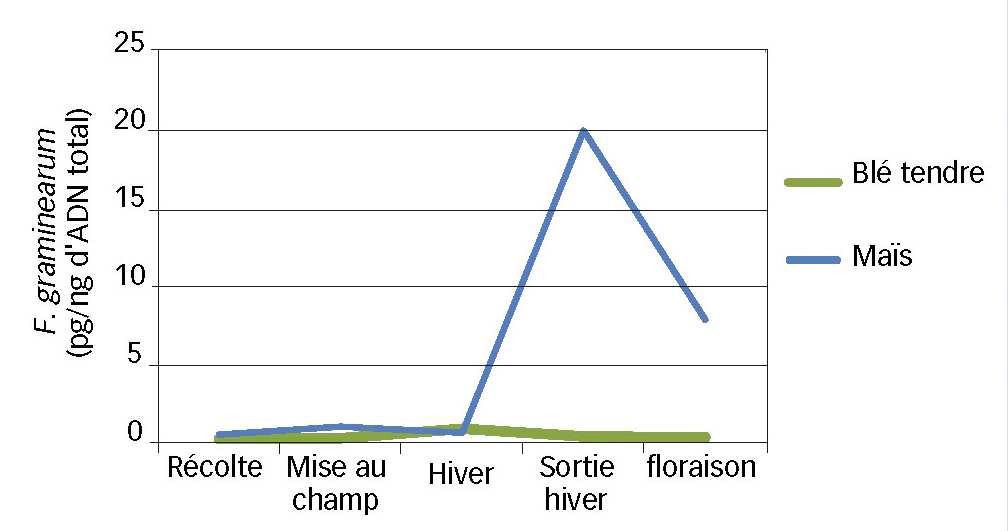
Évolution au cours du temps des quantités de F. graminearum sur résidus de maïs et de blé tendre.
Fig. 5 : Au laboratoire, l'eau, il en faut, mais...
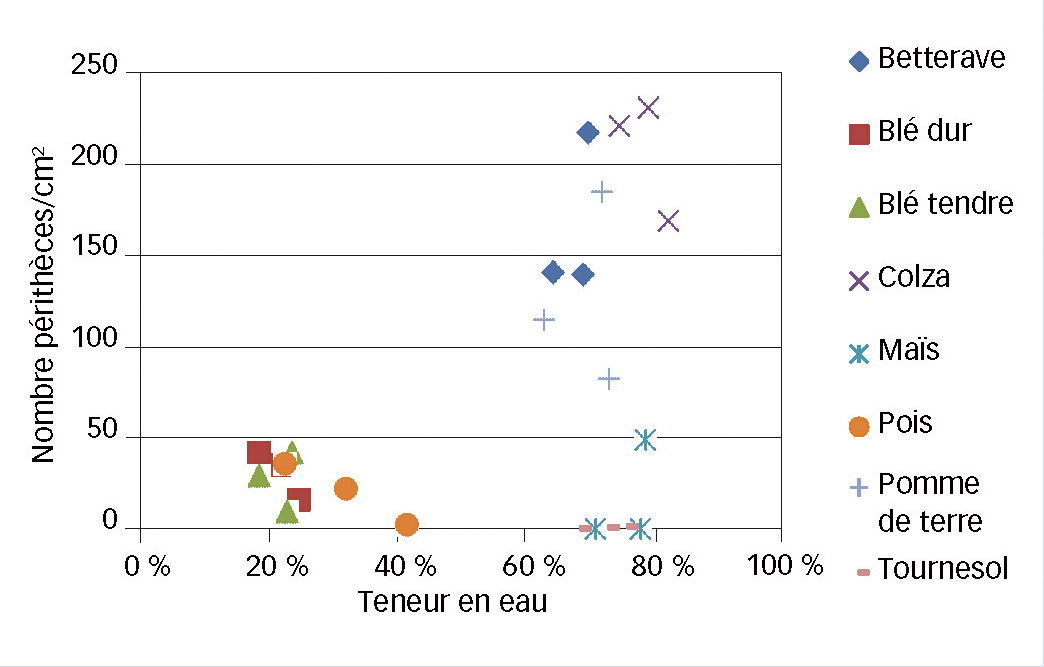
Relation entre teneur en eau des résidus et nombre de périthèces par cm2 : l'eau est nécessaire mais pas suffisante.
Fig. 6 : Au laboratoire, l'azote va avec les périthèces
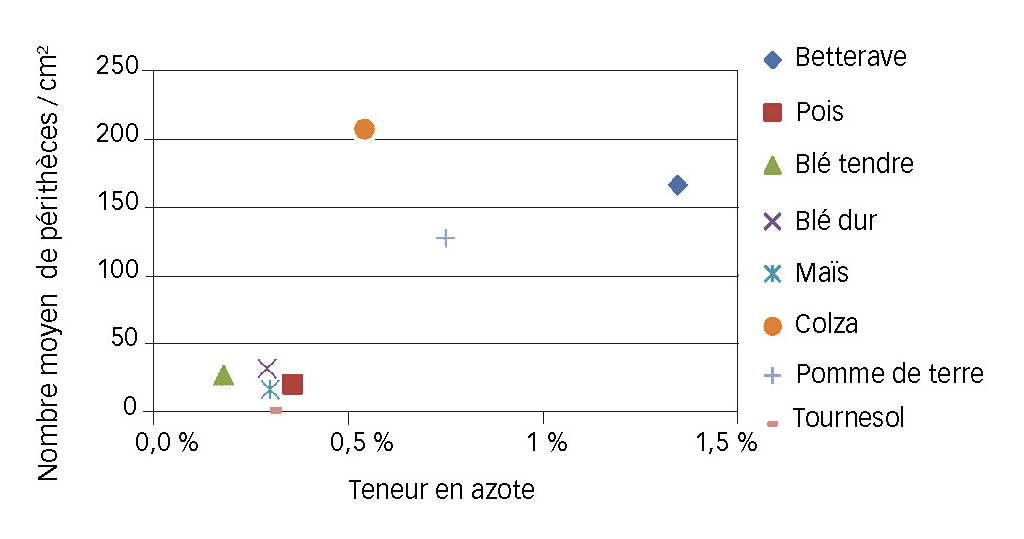
Relation entre teneur en azote des résidus et nombre de périthèces par cm2.
Fig. 7 : Au champ, la nuisibilité, sur blé, des débris de maïs
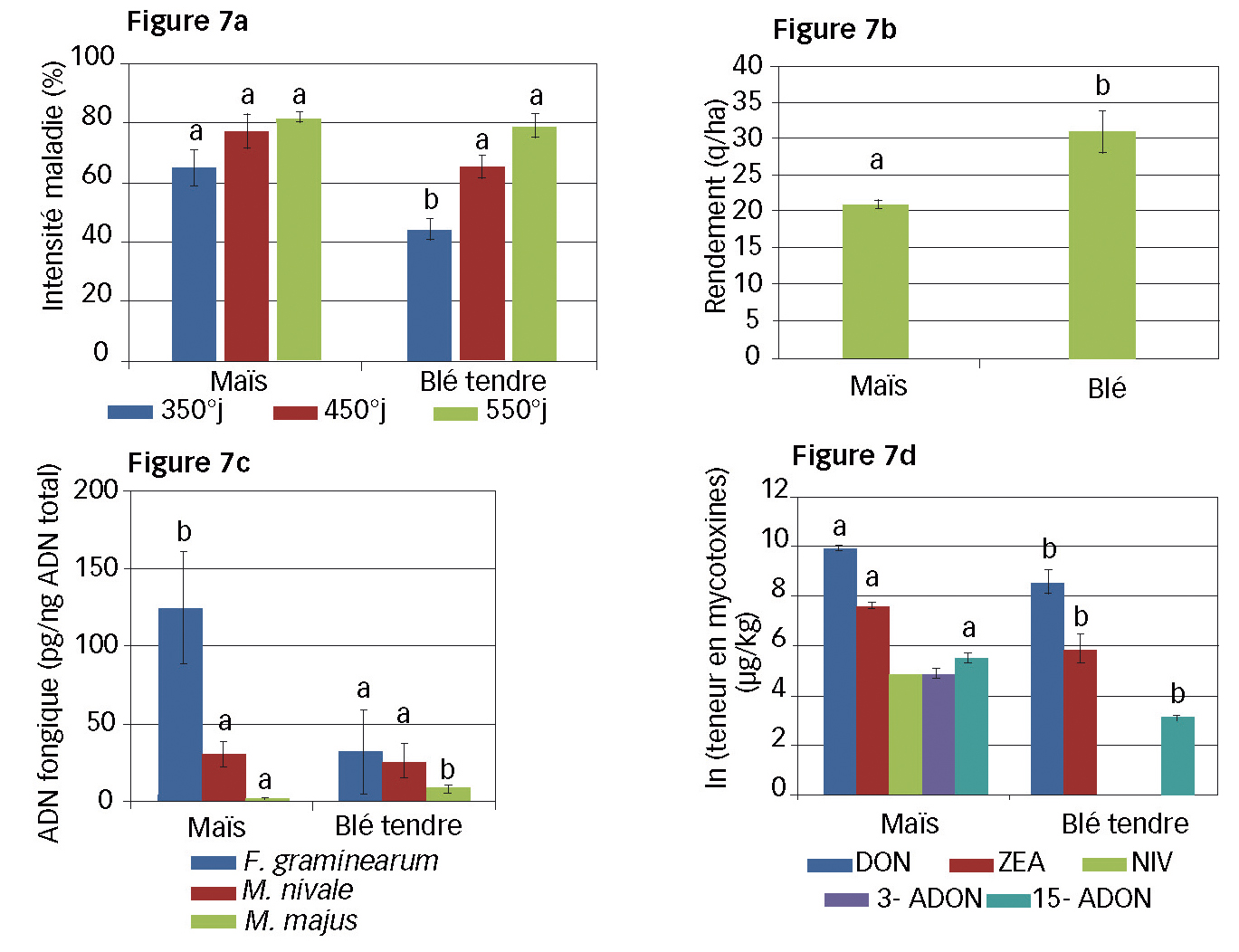
7a : Intensité de la maladie sur épi de blé tendre, selon le type de résidu au sol ;
7b : Rendement (q/ha) des micro-parcelles de blé tendre selon le type de résidu au sol ;
7c : Quantité d'ADN fongique des grains de blé tendre selon le type de résidu au sol ;
7d : Teneurs en mycotoxines (en ln) des grains récoltés selon le type de résidu au sol.
Les lettres a et b indiquent des moyennes significativement différentes (Test de Student).

