 Imprimer
Imprimer dans une parcelle de colza fortement parasitée (Gibot-Leclerc et al., 2012). Photo : S. Gibot-Leclerc")
Hampe florale de Phelipanche ramosa (orobanche rameuse) dans une parcelle de colza fortement parasitée (Gibot-Leclerc et al., 2012). Photo : S. Gibot-Leclerc
L'orobanche rameuse (Phelipanche ramosa (L.) Pomel) (photo ci-contre) est une plante connue pour parasiter préférentiellement la tomate dans les pays du bassin méditerranéen, le tabac et le chanvre en Europe centrale. Mais le colza semble être un nouvel hôte préférentiel avec une augmentation du nombre de parcelles infectées en Bulgarie, Espagne, France et Italie.
Pourquoi cette orobanche inquiète
Une expansion massive sur colza
En France, on observe une expansion massive de cette espèce dans les parcelles de colza depuis la fin des années 1990. La perte de rendement peut aller jusqu'à 90 % dans les cas les plus graves. La culture de colza est même devenue l'hôte principal du parasite avant le chanvre, le tabac et le sarrasin.
Des relais chez les adventices
L'orobanche est capable d'effectuer son cycle de vie complet non seulement sur des espèces cultivées mais aussi sur une cinquantaine d'espèces adventices.
En l'absence de culture hôte, les adventices peuvent prendre le relais pour permettre la reproduction du parasite et accroître son stock de semences dans le sol, facilitant ainsi l'infection des cultures suivantes.
Cette tendance pourrait être renforcée avec les remontées de populations d'adventices liées aux réductions des molécules herbicides autorisées au niveau européen (voir Liens utiles p. 47) et la nécessité de réduire la consommation des pesticides dans le cadre du programme Ecophyto (voir Liens utiles p. 47).
Face à elle, on n'a que les pratiques culturales et l'abandon des cultures hôtes
Actuellement, il n'y a pas d'herbicide homologué pour lutter contre l'orobanche. Son contrôle est principalement fondé sur des méthodes préventives à effet partiel : travail du sol pour limiter le contact entre les racines de l'hôte et les semences du parasite, ou diminution de la fréquence des cultures sensibles dans la rotation. Identifier des combinaisons judicieuses de techniques culturales au sein des systèmes de culture est donc devenu crucial pour contrer l'expansion du parasite.
En Côte-d'Or et en Haute-Saône, l'orobanche rameuse est en pleine expansion. La situation devient de plus en plus préoccupante sur chanvre et colza, contraignant les agriculteurs à changer de cultures sur leurs parcelles parasitées.
C'est dans ce contexte inquiétant, tant au niveau local que national, que nos travaux de recherche sont menés au sein de l'UMR 1347 Agroécologie (AgroSup Dijon-Inra-université de Bourgogne) de Dijon.
Des durées de cycle différentes selon la culture parasitée
L'orobanche rameuse est une espèce autogame, c'est-à-dire fécondée par son propre pollen. Deux traits de vie peuvent avoir une incidence sur ses performances :
(1) la gamme d'hôtes, c'est-à-dire la capacité d'infecter un grand nombre de plantes avec plus ou moins d'efficacité,
(2) l'histoire de vie, c'est-à-dire la capacité de se développer et de produire des graines avant la mort de son hôte.
Selon son hôte, cette orobanche montre des durées variables de cycle. Elle a un cycle de 40 semaines sur colza, mais de 14 à 16 semaines sur tabac, tomate, sarrasin, chanvre et de 7 semaines sur Arabidopsis thaliana (Gibot-Leclerc et al., 2012).
L'orobanche semble donc ajuster la durée de son cycle à celui de son hôte.
Bien que des facteurs environnementaux (température ou photopériode) puissent influer sur sa durée de cycle, il est probable que ce trait soit principalement contrôlé par des interactions hôte-parasite lui permettant de maximiser l'absorption des ressources de l'hôte et son succès reproducteur.
Pourquoi une telle variabilité de durée de cycle ?
Il est essentiel de le comprendre
Afin de développer des stratégies de lutte durables, il est essentiel de comprendre si les différences de durées de cycle observées sur le terrain sont déterminées par l'hôte (synchronisation du cycle du parasite avec celui de son hôte) ou par la propre identité génétique du parasite (pathovars). Il est également important de décrire la variabilité génétique parmi les populations de l'orobanche (Gibot-Leclerc et al., 2013a).
Test d'infections croisées et caractérisation génétique sur deux populations
Notre expérimentation d'infections croisées pour comparer le succès d'infection de deux populations d'orobanche (P-long récoltées sur colza et P-short récoltées sur tabac) sur deux espèces hôtes (le colza, culture à cycle long et la tomate, culture à cycle court), a mis en évidence que la durée du cycle de l'orobanche est principalement déterminée par l'identité de la population du parasite plutôt que par le cycle de l'hôte.
D'autre part, la caractérisation de la variation génétique de ces deux populations à un gène impliqué dans la durée du cycle de vie (gène de floraison FT) a démontré que l'acquisition du nouvel hôte préférentiel (le colza) depuis la fin des années 1990 est associée à une divergence génétique.
Ces résultats nous permettent d'émettre l'hypothèse que le transfert de préférence d'hôte vers le colza (culture à cycle long), se serait accompagné d'une évolution de la durée du cycle du parasite (Figure 1).
Il y aurait eu sélection de génotypes d'orobanche à cycle long et donc acquisition d'une nouvelle niche écologique puisque ces génotypes ne sont plus en mesure de terminer leur cycle de vie sur une culture à cycle court comme la tomate.
Une nouvelle catégorie d'hôtes...
Trois types d'interactions connus
Jusqu'à présent, trois types d'interactions hôte-parasite ont été identifiés pour des couverts monospécifiques (composés d'une seule espèce hôte) :
– la plante hôte : elle stimule la germination du parasite qui se fixe ensuite sur ses racines jusqu'à la production de graines,
– la plante non-hôte : elle ne stimule pas la germination du parasite et n'est donc pas parasitée,
– la plante faux hôte : elle stimule la germination du parasite sans pour autant permettre le développement du parasite.
Étude sous couverts plurispécifiques
Pour des couverts plurispécifiques (composés d'au moins deux espèces), la germination et la fixation de l'orobanche n'ont jamais été étudiées. Or des associations plurispécifiques sont fréquemment utilisées, en conditions tropicales, pour protéger les cultures hôtes sensibles à l'orobanche avec des espèces non-hôtes.
Nous avons donc décidé d'étudier la germination du parasite et sa fixation dans des couverts monospécifiques vs. plurispécifiques, associant l'orobanche et une plante hôte (le colza), une plante non-hôte (le liseron des champs Convolvulus arvensis L.) ou une combinaison des deux (Gibot-Leclerc et al., 2013b).
Surprise, le liseron facilitateur
Contre toute attente, nous avons observé une multiplication de l'infection du colza par environ trois en présence du liseron des champs, comparé au colza seul. Vis-à-vis de l'orobanche rameuse, le liseron des champs est classé comme espèce non-hôte.
Ce résultat significatif nous a conduits à repenser la classification actuelle des espèces en fonction de leur sensibilité à l'orobanche. Nous proposons maintenant une nouvelle catégorie d'espèces de plante appelée « non-hôte facilitateur ».
... découverte de la facilitation
Un nouveau processus d'interaction
Nous avons ainsi mis en évidence un nouveau processus d'interaction hôte-parasite chez l'orobanche, appelé « facilitation », au cours duquel l'infection du colza (plante hôte) par l'orobanche augmente quand le liseron (plante non-hôte) se situe à proximité.
Parallèlement, nous avons observé que le liseron des champs est capable de supporter des fixations secondaires du parasite issues des fixations primaires observées sur les racines du colza situé à proximité. Le mécanisme sous-jacent demeure encore inconnu.
Repenser la gestion du parasite
Introduire des cultures non-hôtes... à condition qu'elles ne soient pas facilitatrices !
Notre découverte implique de repenser la gestion de l'orobanche dans les cultures. La rotation des cultures avec des cultures non-hôtes est une recommandation générale pour les agriculteurs mais nos résultats indiquent que cette stratégie serait peut-être beaucoup moins efficace que prévu. Certaines cultures non-hôtes pourraient être des non-hôtes facilitateurs.
De même, des espèces adventices identifiées comme non-hôtes pourraient augmenter l'infection de plantes hôtes par le processus de facilitation. Il faut donc évaluer à la fois la culture et la flore adventice associée pour leur capacité à déclencher et/ou fixer le parasite, et ce, tant en couverts monospécifiques que plurispécifiques.
Besoin de modèles
Par ailleurs, comme les graines d'orobanche peuvent survivre dans le sol pendant plusieurs années, le contrôle du parasite ne peut pas être simplement évalué à l'échelle annuelle mais doit être optimisé à l'échelle pluriannuelle.
En raison de cet effet à long terme et de la multiplicité des techniques culturales qui influent potentiellement sur la dynamique du parasite, soit directement, soit via la flore adventice, le développement de modèles permettant de quantifier l'effet des systèmes de culture sur la dynamique du parasite en interaction avec les adventices non-parasites devient crucial.
Un projet de modélisation pour lutter contre l'orobanche
Objectif et version préliminaire
Notre objectif est de développer un modèle mécaniste, appelé PHERASYS, quantifiant les effets de système de culture sur la dynamique du parasite, en interaction avec la flore adventice non-parasite, et de le coupler avec le modèle existant, FLORSYS, qui prédit la flore adventice dans les systèmes de culture (Gardarin et al., 2012).
Le modèle couplé FLORSYS-PHERASYS est un ensemble de deux cycles de vie, le premier pour les adventices non-parasites annuelles (dorénavant « adventices ») et la culture, et le second pour l'orobanche (dorénavant « parasite ») (Figure 2). Nous avons développé une version préliminaire de PHERASYS (Colbach et al., 2011), à partir de données issues de la littérature, que nous avons utilisée pour tester une méthode d'évaluation de systèmes de culture existants et prospectifs pour le risque d'orobanche et la contribution de la flore adventice à la dynamique du parasite.
Trois expérimentations en cours
Cette version préliminaire a permis d'identifier les lacunes de connaissances sur le cycle de vie du parasite et ses interactions avec les adventices non-parasites, ce qui nous a amenés à planifier trois expérimentations :
– la quantification du potentiel infectieux des semences du parasite en fonction de leur âge et de la saison,
– l'analyse de l'architecture racinaire des plantes cultivées et adventices pour prédire la proximité entre les semences du parasite et les racines de l'hôte et donc la proportion de graines du parasite pouvant germer puis se fixer sur un hôte,
– enfin, l'étude des relations trophiques entre l'hôte et le parasite pour prédire la biomasse accumulée par le parasite au dépend de l'hôte.
Ces expérimentations sont actuellement en cours afin de développer la nouvelle version de PHERASYS.
Des simulations encourageantes mais pas encore de conseils prêts à l'emploi
Les simulations confirment les risques liés aux adventices hôtes
Nous avons déjà réalisé quelques simulations de systèmes de culture avec la version préliminaire du modèle FLORSYS-PHERASYS dans le but d'évaluer des systèmes de culture existants et prospectifs pour leur risque d'orobanche et la contribution de la flore adventice à l'infection par le parasite.
Le système de référence est une rotation typique du nord-est de la France, colza-blé d'hiver-orge d'hiver sans labour. La présence d'une flore adventice contenant des espèces hôtes augmente le stock semencier du parasite avant le semis du colza d'environ 70 %, toutes modalités confondues (Figure 3).
Des pratiques qui font augmenter ou baisser le risque
Introduire du pois avant le colza dans la rotation diminue le risque parasitaire de plus de 90 %, et le surplus de semences du parasite dû aux adventices devient alors négligeable.
Labourer après colza a peu d'influence directe sur le parasite mais diminue le risque via les adventices de plus de 60 %. Le semis direct divise le risque parasitaire direct par environ 1 800 parce que les semences du parasite restent majoritairement à la surface du sol, loin des racines. Mais comme l'absence de travail du sol favorise simultanément la flore non-parasite, le risque global est au final multiplié par 5.
Diminuer la densité de semis du colza réduit le risque direct de 30 % en diminuant la probabilité de contact parasite-racine, mais cette pratique laisse davantage de place aux adventices. Au final, elle n'a donc que peu d'impact.
Retarder le semis des cultures de trois semaines diminue les risques directs et indirects et réduit le stock global du parasite d'environ 30 %.
Prudence, ce ne sont que des illustrations !
Bien que cohérents avec les connaissances actuelles sur le parasite, ces résultats de simulation ne sont qu'une illustration des applications possibles du modèle et non pas des conseils prêts à l'emploi. En effet, PHERASYS doit d'abord être amélioré puis évalué avec des données indépendantes de terrain pour déterminer son domaine de validité.
Fig. 1 : Deux populations comparées en test d'infections croisées
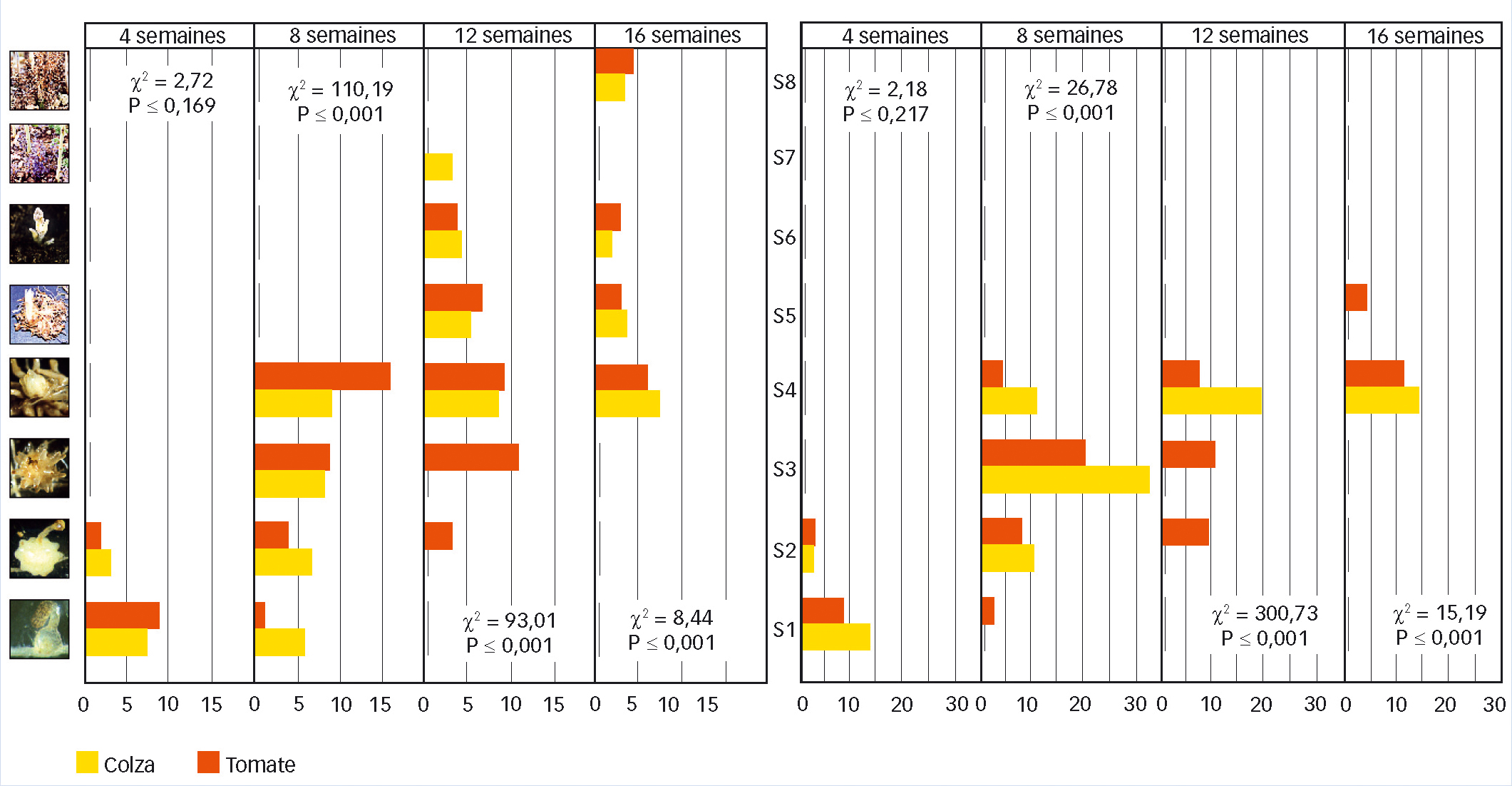
Distribution des stades de développement de deux populations d'orobanche dites P-short (à gauche) et P-long (à droite) sur le colza (jaune) et la tomate (rouge). Valeur (χ2) et probabilité (P) du test du Chi-deux.
Les valeurs sur l'axe des abscisses correspondent aux racines carrées du nombre d'individus observés. Note : les échelles ne sont pas les mêmes sur les deux figures. Les stades sont, de bas en haut : S1 : fixation, S2 : jeune tubercule, S3 : tubercule âgé, S4 : bourgeon, S5 : tige souterraine, S6 : émergence, S7 : floraison et S8 : fructification.
Cette figure montre la dynamique d'infection de chaque population sur chaque hôte. Malgré son plus faible succès d'infection pendant toute la durée de l'expérimentation, seul P-short termine son cycle et produit les stades reproducteurs sur les deux hôtes (tomate et colza). Inversement, P-long n'a pas pu terminer son cycle pendant la durée de l'expérimentation, et ce, quel que soit l'hôte.
L'analyse des résultats suggère que P-long possède un taux de mortalité élevé au stade bourgeon sur son hôte naturel (colza), mortalité pouvant être causée par des mécanismes de résistance.
Fig. 2 : Vue sur le modèle
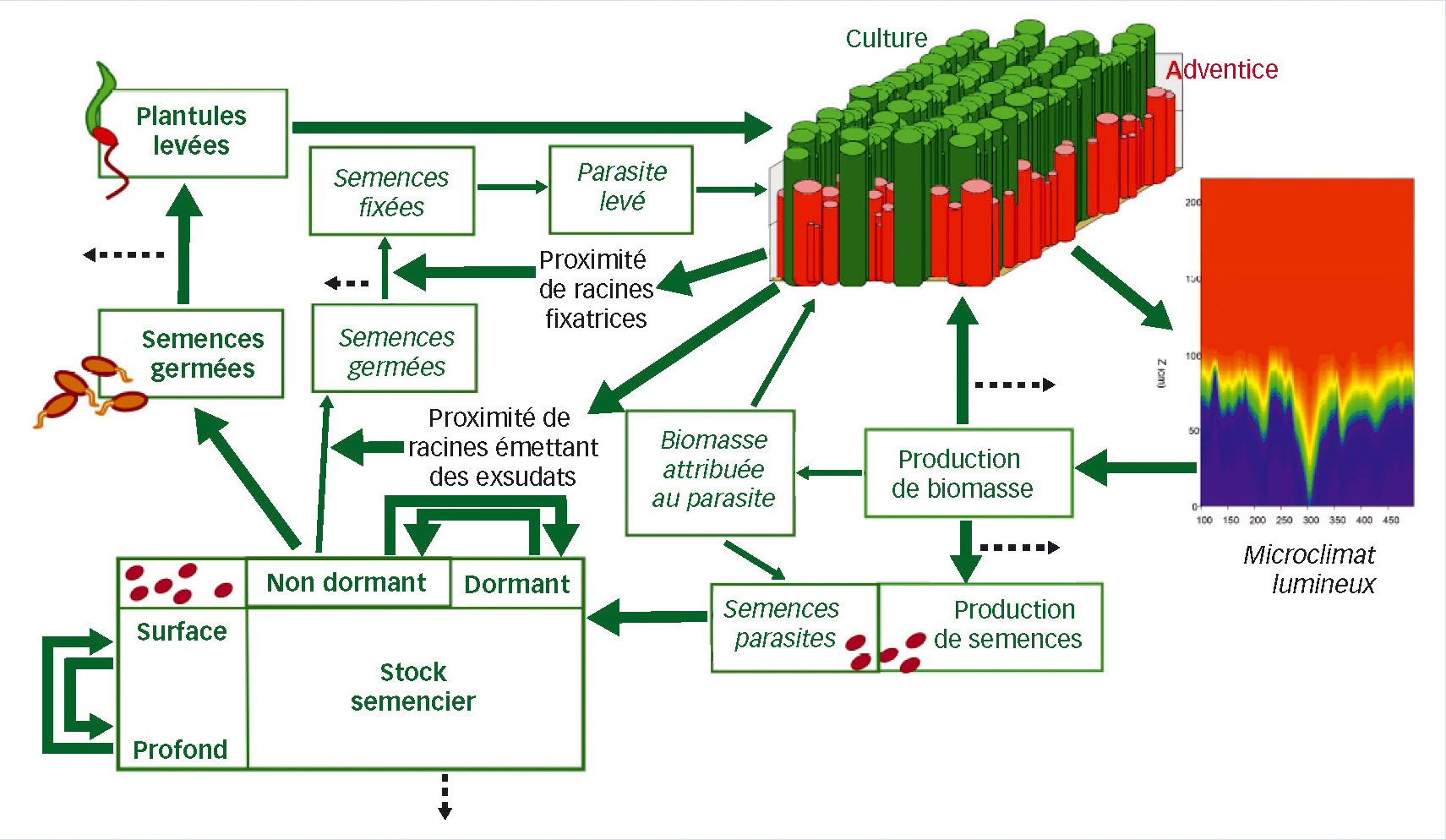
Organigramme du modèle couplé FLORSYS-PHERASYS prédisant la dynamique de l'orobanche et des adventices non parasites dans les systèmes de culture. Cycle de vie représentant les stades des adventices non-parasites (plantules levées, ←) et parasites (semences fixées →), avec des processus de mortalité (← - - -).
Fig. 3 : Simulation de la stimulation, ou non
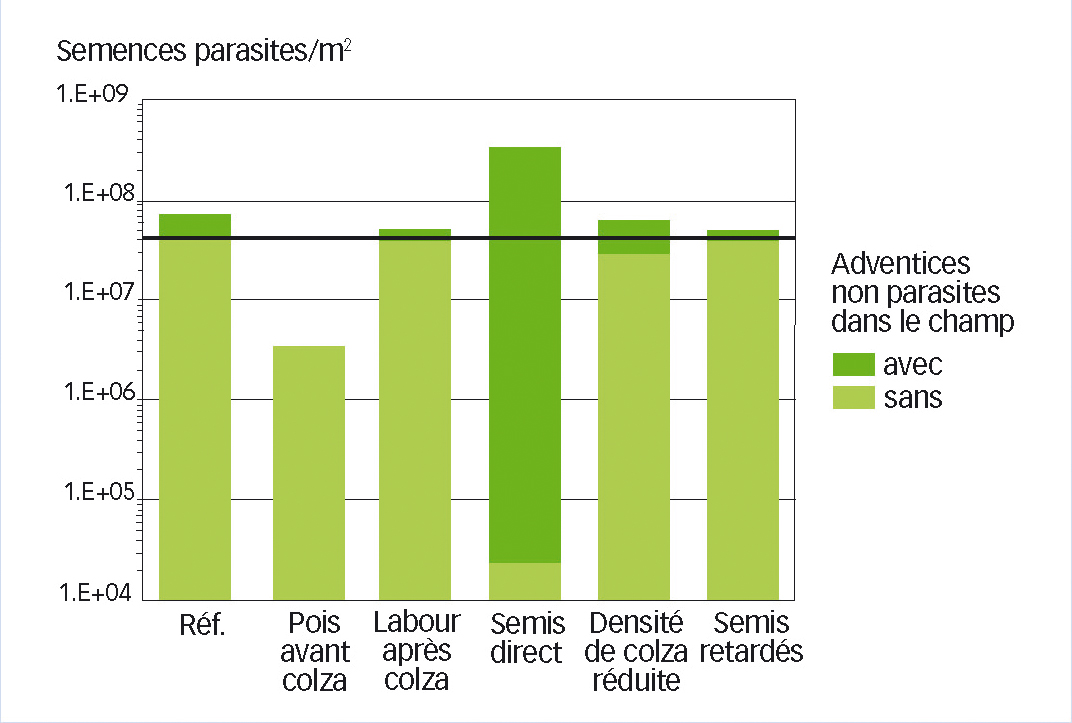
Impact du système de culture et de la flore adventice non-parasite sur le risque d'infection du parasite dans une rotation colza/blé/orge. Stock semencier de l'orobanche dans le sol avant colza après 12 années de simulation avec PHERASYS (Colbach et al., 2011).

