 Imprimer
Imprimer et bleuet (Cyanus segetum), deux espèces caractéristiques du blé d'hiver. La première est restée commune, la seconde est devenue rare dans la plupart des régions. Photo : G. Fried")
Coquelicot (Papaver rhoeas) et bleuet (Cyanus segetum), deux espèces caractéristiques du blé d'hiver. La première est restée commune, la seconde est devenue rare dans la plupart des régions. Photo : G. Fried

Miroir-de-Vénus L. speculumveneris, messicole rare.

Véronique à feuilles de lierre V. hederifolia, restée commune. Photos : G. Fried
Dans la précédente édition de Phytoma(1), était exposée l'analyse statistique des changements de fréquence des espèces adventices du blé d'hiver au cours des années 2000, montrant des évolutions modérées parmi les espèces les plus communes.
Mais ce résultat masque en réalité de profondes disparités régionales. Nous allons chercher à les expliquer dans ce second article.
Un objectif et deux volets
Savoir comment les pratiques influencent la flore
Le réseau Biovigilance Flore nous fournit des données récoltées sur plusieurs centaines de parcelles, où la flore observée, les pratiques agricoles et les conditions environnementales sont enregistrées simultanément.
Cet article utilise ces données pour analyser comment les pratiques agricoles influencent la flore du blé, vu le challenge actuel de leur gestion : maintenir la production agricole et conserver la biodiversité adventice à la base de chaîne trophique dans l'agroécosystème (Marshall et al., 2003 ; Storkey 2006). Dans une première partie, nous évaluons le niveau de contrôle exercé par un itinéraire technique sur les espèces adventices (en distinguant espèces communes et espèces messicoles rares) et sur la diversité de la flore qui en résulte.
Dans une seconde partie, afin de mieux comprendre les règles sous-jacentes aux variations de la diversité floristique observée, nous nous intéressons aux mécanismes de sélection de la flore en analysant les caractéristiques biologiques des espèces corrélées aux pratiques culturales et au statut en progression et en régression.
Le travail réalisé
Base de données sur 953 parcelles
Parmi les 953 parcelles du réseau Biovigilance Flore utilisées pour mesurer les changements floristiques(1), plusieurs sous-ensembles différents du jeu de données ont été constitués selon les questions à aborder et la disponibilité des données agricoles et environnementales.
Indice de contrôle de la flore dans 516 relevés
Nous avons tiré profit des quatre notations (zones « témoin » et « désherbée » en automne et au printemps) pour calculer un indice brut de contrôle pour chaque espèce (IC, voir formule page suivante) et chaque parcelle (ICpar, voir Fried et al., 2013 pour plus de détails).
Cet indice mesure la différence de densité entre la dernière notation dans la zone désherbée et les autres notations (zones témoins et traitées en automne (N1) et zone témoin au printemps (N2 tem) prises ensemble comme ayant valeur de contrôle. Pour ce calcul, nous n'avons conservé que les 516 relevés où la notation N2 avait été réalisée au moins 10 jours après le dernier désherbage afin de capturer au mieux l'effet de l'ensemble des traitements.
Facteurs de diversité floristique évalués
Les facteurs susceptibles de faire varier le nombre d'espèces par parcelle ont été recherchés à trois niveaux :
1) au niveau des relevés, en distinguant les quatre notations annuelles afin de mesurer l'influence brute du traitement herbicide (zones témoin/traitée) et de la date (N1/N2) ;
2) à l'échelle du relevé synthétique de parcelle (toutes les espèces identifiées dans les quatre notations d'une campagne n) afin de mesurer l'influence relative pratiques culturales/ conditions environnementales ;
3) aux échelles communale et départementale (moyenne des 935 parcelles au sein de 438 communes et 57 départements) en lien avec l'indicateur des systèmes agricoles à haute valeur naturelle (HVN) bâti à l'échelle communale (Pointereau et al., 2010). L'indice HVN évalue le degré d'intensification agricole sur la base de la diversité d'assolement, de l'extensivité des pratiques (source : enquêtes Agreste) et de la densité des éléments paysagers. Selon une précédente étude (Doxa et al., 2010), cet indice est bien corrélé à l'abondance et à l'indice de spécialisation des oiseaux des milieux cultivés.
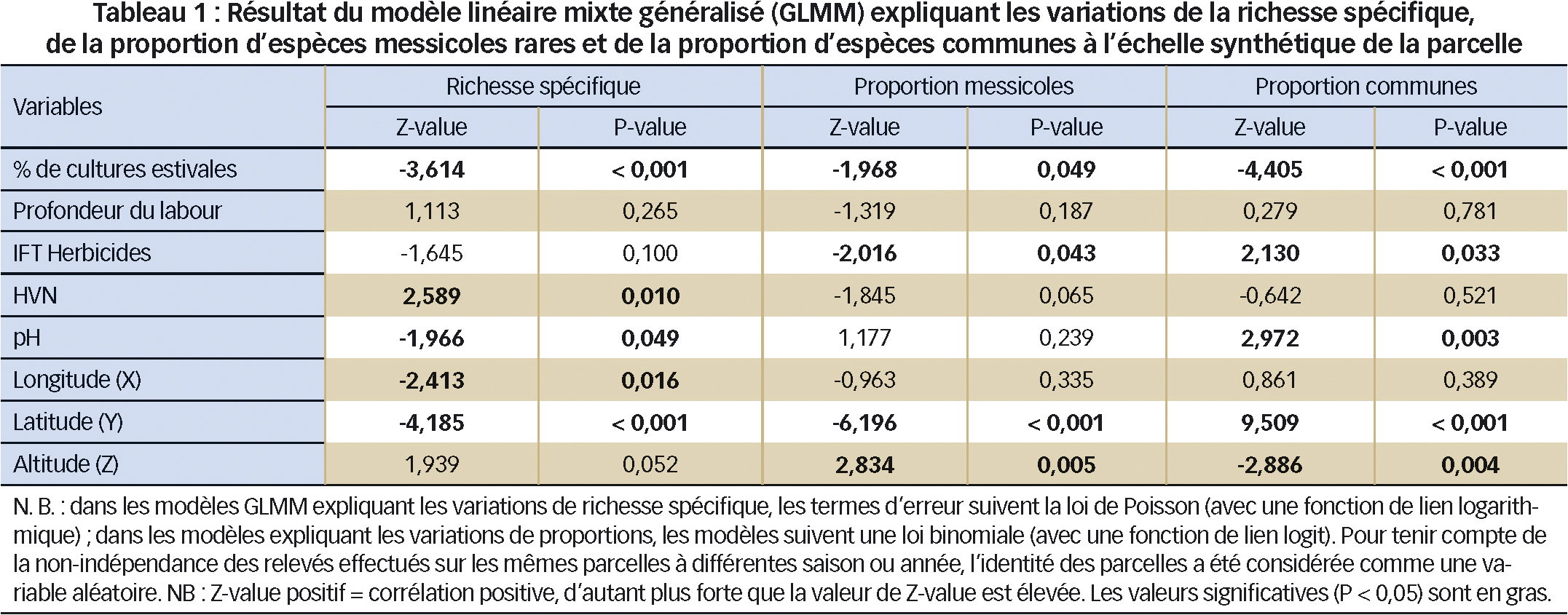
Pour modéliser les variations de diversité floristique, des modèles linéaires généralisés mixtes (GLMM) ont été utilisés à l'échelle des 5 080 relevés (4 notations x 1 270 parcelles) ou de 890 parcelles. Les variables explicatives incluent la campagne culturale (2003 à 2010), la région (Est, Massif central, Nord-Bassin parisien, Ouest, Sud-Est, Sud-Ouest), la date (N1, N2) et la zone à l'intérieur de la parcelle (Témoin, Traitée). À l'échelle de la parcelle, les variables explicatives sont la latitude (Y), la longitude (X), l'altitude (Z), le pH, l'indice de fréquence des traitements (IFT) en herbicides, la profondeur maximale de labour, la proportion de cultures estivales dans la rotation et l'indice HVN (Tableau 1).
Suivant les cas, la variable à expliquer est « S », la richesse spécifique totale, « Smess », celle en espèces messicoles rares, « Pmess », la proportion en espèces messicoles rares et « Pcom », celle en espèces communes (= présentes dans plus de 15 % des relevés, N = 18 espèces sur 261 espèces détectées).
Enfin, nous avons testé la corrélation entre l'indice HVN et la richesse spécifique S et Smess moyennée à l'échelle communale.
Lien flore/pratiques dans 218 parcelles
Le lien entre la flore et les pratiques a été analysé en reliant directement les caractéristiques biologiques (traits) des espèces (dates de levée et floraison, hauteur, poids des graines, etc.), les conditions pédoclimatiques et les pratiques agricoles grâce à une analyse multivariée dite RLQ (détails dans Fried et al., 2012). Dix traits ont été croisés à 10 variables agronomiques et environnementales sur un échantillon de 218 parcelles de blé d'hiver de la période 2003-2006. L'analyse regroupe les espèces en fonction de leurs traits et de leur réponse aux variables, permettant ainsi d'identifier les combinaisons « traits x pratiques » associées à des progressions ou régressions d'espèces.
Attention : cette approche basée sur des corrélations ne permet pas d'affirmer des liens de causalité.
Quels facteurs déterminent le nombre d'espèces dans les parcelles de blé ?
Le blé : culture à faible diversité parcellaire, avec des influences régionale et pédoclimatique
Le nombre moyen d'espèces par parcelle de blé d'hiver est de 10,51 (min = 1, max = 34). À l'inverse d'une idée reçue, le blé est une culture à faible diversité parcellaire en comparaison du colza (12,57, min = 1, max = 37), du maïs (14,94, min = 1, max = 45) et du tournesol (16,2, min = 1, max = 53).
Les résultats du modèle GLMM (Tableau 1) indiquent que le nombre d'espèces dans les champs de blé varie fortement en fonction de la latitude (Y) et de la longitude (X). La Figure 1 et la Figure 2A illustrent ce contraste entre zones de forte diversité (Massif central, Ouest) et celles plus faible (Bassin parisien, Nord et Est).
La proportion d'espèces messicoles répond fortement à la latitude (z = -6,196, P < 0,001, Tableau 1) et à l'altitude, (z = 2,834, P = 0,005) : elle est plus élevée dans le sud de la France ainsi qu'aux étages collinéen et montagnard inférieur. Le type de sol joue aussi avec des parcelles plus riches sur sol à pH acide (z = -1,966, P < 0,049).
La relation négative entre pH et diversité de la flore peut résulter de pratiques peu intensives sur sols acides, souvent pauvres et filtrants. L'augmentation de la richesse en espèces des pôles vers l'équateur est un fait observé pour la plupart des groupes taxinomiques. Enfin, nombre de messicoles sont des espèces xéro-thermophiles et calcicoles : elles trouvent des conditions propices à leur développement dans le Midi.
Mais les effets géographique et altitudinal pourraient aussi être liés aux systèmes de culture, avec des parcelles plus riches en zones de polyculture-élevage à pratiques extensives. C'est ce que suggère l'analyse de la relation flore/indice HVN.
Influence des pratiques agricoles
En moyennant la diversité de la flore du blé par commune (n = 438 communes), on note une corrélation positive avec l'indice de système agricole à « haute valeur naturelle » (HVN) reflétant l'extensivité des pratiques et la diversité des paysages (test de corrélation de Spearman, rho = 0,133, P = 0,005). La tendance est identique pour la richesse en espèces messicoles rares (rho = 0,095, P = 0,047). S'il existe une large gamme de niveaux de richesse floristique sur des communes à valeurs d'indice HVN faibles, toutes les communes à indice HVN élevé (> 15) ont des parcelles relativement riches. Le seuil de 14,78 points est d'ailleurs celui au-delà duquel une commune est classée comme ayant une agriculture HVN.
Il reste à déterminer les parts de diversité expliquées par la diversité du paysage (Gaba et al., 2010), celle des assolements ou l'intensité des pratiques. Les données du réseau Biovigilance fournissent une première réponse (Tableau 1) :
– la profondeur du labour n'a pas d'influence significative ;
– en revanche, l'augmentation de l'IFT réduit la proportion de messicoles et favorise celle des espèces les plus communes. Cela renforce l'impression déjà signalée (voir note de bas de page p. 43) que la sensibilité aux herbicides constitue un facteur actif du turnover observé au sein des principales espèces ;
– le pourcentage de cultures estivales dans les précédents du blé est corrélé négativement à la richesse spécifique. Cet effet serait surtout lié aux blés semés après une longue succession de cultures estivales (proportion > 0,8) à diversité faible, notamment des espèces caractéristiques du blé.
On peut donc opposer les messicoles, surtout présentes en zones d'agriculture extensive de moyenne montagne « méridionale », type Lubéron, où beaucoup d'espèces ont trouvé un dernier refuge (Jauzein, 2001), et les espèces communes davantage représentées en systèmes intensifs de plaine céréalière du Nord.
Indice de contrôle de la flore et herbicides
Toutes régions et notations confondues (5 080 relevés), le nombre moyen d'espèces par relevé dans le blé d'hiver est de 5,25 (minimum 0, max. 32, 1er quartile = 2, 3e quartile = 7). Les variations sont liées à l'effet des herbicides (différence placettes témoin/traité, z = 11,26, P < 0,001) et à un effet date de notation (différence entre N2 et N1, z = 9,93, P < 0,001) avec interaction « Date »/« Herbicide » significative (z = 7,43, P > 0,001). Le nombre moyen d'espèces par relevé (Figure 2 B) est minimal en automne dans les parties traitées (moyenne 4,02 ± 3,72 espèces) et maximal en zone témoin au printemps (7,19 ± 4,75 espèces) quand les levées d'espèces printanières complètent la flore à germination automnale. En comparant la zone désherbée après désherbage aux autres notations, l'indice moyen de contrôle par parcelle (ICpar) s'élève à 7,1 (au moins sept espèces éliminées), soit un peu moins que la valeur trouvée en colza (8,8 : Fried et al., 2013). C'est davantage que la différence moyenne du nombre d'espèces entre zones témoin et traitée (environ trois espèces, Figure 2B) mais plus proche de la réalité. En effet, une espèce passant d'un score d'abondance de 5 à 1 passe d'une densité de 75,5 ind/m² à 0,5 ind/m² pour une richesse spécifique inchangée (présente qu'elle soit à 0,5 ou 75,5 ind./m²).
Au niveau des espèces, le faible niveau de contrôle de la folle avoine (Avena fatua, IC = 0,30, signifiant peu de différences de densité entre zone traitée et témoin) corrobore la progression récente de cette espèce plutôt printanière(1). Enfin, si le vulpin (Alopecurus myosuroides, IC = 0,69) est assez bien contrôlé (sauf populations résistantes) ainsi que le pâturin annuel (Poa annua, IC = 0,69), en revanche les ray-grass (Lolium spp., IC = 0,50), le brome stérile (Bromus sterilis, IC = 0,37), et surtout la vulpie queuederat (Vulpia myuros, IC = 0,14) sont assez mal maîtrisés... donc à surveiller !
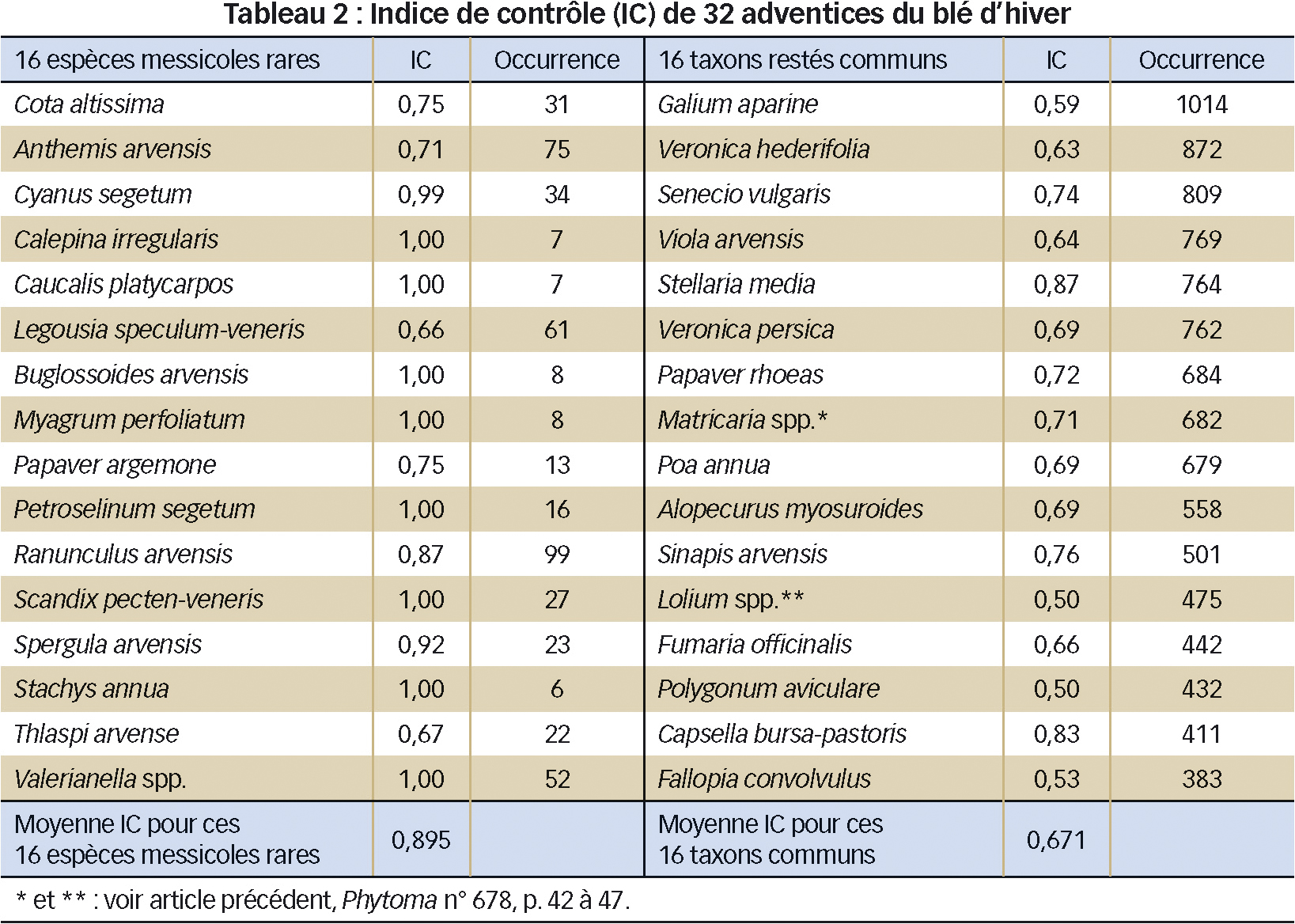
Selon le Tableau 2, les seize espèces messicoles rares présentes dans au moins cinq relevés ont une baisse de densité significativement plus marquée (IC = 0,90 +/0,14) que les seize taxons (espèces ou groupes d'espèces) les plus communs (IC = 0,67 +/0,11) entre zone témoin et zone désherbée (test de Mann-Withney, U = 30,5, P < 0,001).
Au-delà de la corrélation entre intensité du désherbage (IFT) et certains groupes d'espèces, établie à l'échelle de la parcelle, le dispositif semi-expérimental de placettes témoin/traité illustre plus directement l'effet des herbicides sur les diverses espèces. Cela nous mène à la dernière partie qui vise à mieux identifier quelles espèces sont sélectionnées par les différentes pratiques.
À retenir : réponse de la flore aux pratiques
Influence du travail du sol
L'analyse multivariée RLQ montre une corrélation significative entre les variations des pratiques et de l'environnement d'une part et des traits des adventices d'autre part (Figure 3).
Concernant le travail du sol, trois tendances ressortent : 1/ Le non-travail du sol est associé à des espèces pluriannuelles passant la mauvaise saison sous forme de rosettes (hémicryptophytes à rosette) et dispersées par le vent : pissenlit (Taraxacum sect. Ruderalia), picris fausse vipérine (Helminthotheca echioides), picris fausse-épervière (Picris hieracioides) et épilobe à quatre angles (Epilobium tetragonum) (groupe 7 et en partie 6, en bas de la Figure 3).
2/ Le travail réduit (type TCS) semble plutôt favoriser les vivaces à organes de réserve souterrains (géophytes), notamment celles à racines traçantes et drageons type chardon des champs (Cirsium arvense), ou à rhizomes type chiendent rampant (Elytrigia repens) (groupe 6, Figure 3). Ces espèces peuvent être favorisées par une perturbation légère et mieux contrôlées par des labours plus profonds.
3/ Le travail du sol conventionnel avec labours favorise plutôt les espèces annuelles de petite taille et petites graines : lamier pourpre (Lamium purpureum), stellaire intermédiaire (Stellaria media), etc. à gauche sur la Figure 3.
Date de semis et nature du précédent
On note une réponse marquée des flores du blé aux dates de semis (mi-septembre à fin décembre). Les dates les plus tardives, souvent liées à des précédents récoltés tard (betterave, maïs, tournesol), semblent limiter la présence des espèces à pic de germination précoce à l'automne (vulpin des champs, ray-grass). Les adventices les plus compétitives du blé (folleavoine, gaillet gratteron, ray-grass, vulpin) sont associées à des précédents « céréales d'hiver » ou « colza ». À l'opposé, les précédents « maïs » et probablement les rotations courtes « maïsblé » favorisent les espèces à large plage de germination : pâturin annuel, stellaire intermédiaire, séneçon vulgaire (groupe 1), capables de produire des graines dans les deux cultures.
Pression herbicide
Enfin, mais de façon moins nette, une pression herbicide accrue et diversifiée (variable « nombre de molécules de différentes familles HRAC ») favorise des espèces à germination plus tardive. Ce trait peut conférer la capacité d'échapper aux interventions majeures de désherbage chimique mais aussi mécanique.
Ce que la relation flore-pratiques dit des changements de flore
D'abord, la pluralité des facteurs
Premier constat, la distribution des espèces favorisées au cours des trente dernières années dans l'espace multivarié RLQ des traits et des conditions agronomiques et écologiques (en rouge sur la Figure 3) illustre la pluralité des facteurs impliqués dans les changements floristiques observés.
Ainsi le pissenlit, qui a pu profiter de l'extension du non-labour (lien direct), a peu de choses en commun avec le géranium disséqué qui, associé au précédent colza où il est très favorisé, a « débordé » sur d'autres cultures de la rotation dont le blé, du fait de stocks de semences importants (lien indirect).
Prime aux espèces « petites précoces généralistes »
Dans un deuxième temps, si on compare sur la Figure 3 le groupe fonctionnel 1 qui concentre le plus d'espèces en progression à celui regroupant les espèces en régression (groupe 4), une tendance plus générale se dégage.
Les espèces sélectionnées depuis trente ans ont une plage de germination et de floraison plus longue que la moyenne : elles sont qualifiées de généralistes. Elles peuvent boucler leur cycle, de la germination à la production de semences, dans un grand nombre de cultures et d'intercultures de la rotation. Grâce à ces germinations étalées dans le temps permettant l'existence de plusieurs cohortes, elles peuvent échapper aux périodes de traitements. La clé du succès se résume par un appareil végétatif de petite taille (32,14 cm ± 5,67), des petites graines (0,38 mg ± 0,25) et une floraison précoce dès la sortie d'hiver. Ce groupe 1 comprend des espèces rudérales : pâturin annuel, séneçon vulgaire, lamier pourpre, stellaire intermédiaire, véronique des champs (Veronica arvensis), capselle bourse-àpasteur (Capsella bursa-pastoris).
À l'opposé, les espèces en déclin ont des graines plus lourdes (2,47 mg ± 4,09), une hauteur plus importante (52,53 cm ± 25,76) et une floraison plus tardive en avril-mai. Ce groupe 4 inclut la plupart des messicoles en déclin (miroir-de-Vénus, renoncule des champs, anthémis des champs, etc.). Une étude anglaise (Storkey et al., 2010) aboutit au même constat.
S'il est dans la nature du champ cultivé d'être perturbé, le succès des espèces du groupe 1 peut être interprété comme une réponse à l'intensification des perturbations liées aux pratiques agricoles depuis les années 1970 (travaux mécaniques puissants, déchaumage systématique juste après récolte, traitements herbicides). Petite taille et floraison précoce assurent une production de graines entre deux perturbations en cours de campagne. À l'opposé, les espèces à grosses graines ont généralement un stock de graines plus limité et une dormance plus faible conduisant à un épuisement plus rapide de leur banque de graines face au désherbage intensif.
Conclusions
Les dispositifs de suivi de la flore permettent d'établir des corrélations à grande échelle entre la flore observée et les pratiques agricoles. Si cette échelle de perception fait ressortir l'influence de grands gradients écologiques, on note que certaines pratiques semblent plutôt affecter la composition des espèces présentes sur la parcelle (mode de travail du sol), d'autres touchant plus à la diversité (herbicides) ou influençant simultanément ces deux aspects des communautés floristiques (succession culturale et conditions environnementales). L'utilisation des traits s'est révélée fructueuse pour mieux comprendre la sélection de divers groupes d'espèces selon les pratiques agricoles. Dans le cadre des missions de surveillance biologique du territoire, le suivi dans le temps du niveau de contrôle moyen des espèces via la comparaison des zones témoin et traitée pourrait permettre d'identifier de façon précoce des situations de perte de contrôle d'espèces problématiques, et de chercher dans quelles situations elles sont mieux contrôlées. À l'inverse, cela pourrait aussi détecter les prémices de l'effondrement d'espèces comme les messicoles qui semblent poursuivre leur déclin.
La corrélation entre diversité de la flore et indice HVN illustre la possibilité de maintenir une biodiversité élevée dans des zones adéquates en jouant sur les paramètres de l'indice HVN (intrants, diversité de rotation, hétérogénéité du paysage).
Les réseaux d'observation jouent un rôle primordial dans l'acquisition de connaissances nécessaires à la conception de systèmes de culture permettant d'atteindre des objectifs de production agricole et de maintien de la biodiversité dans le cadre de l'agroécologie.
<p>(1) Voir « Flore du blé d'hiver, quarante ans d'évolution », de G. Fried <i>et al., Phytoma</i> n° 678 - novembre 2014, p. 42 à 47.</p>
Comment est calculé l'IC

Fig. 1 : Cartes départementales de la richesse spécifique moyenne (S) et du nombre moyen de messicoles (Smess) à l'échelle de la parcelle
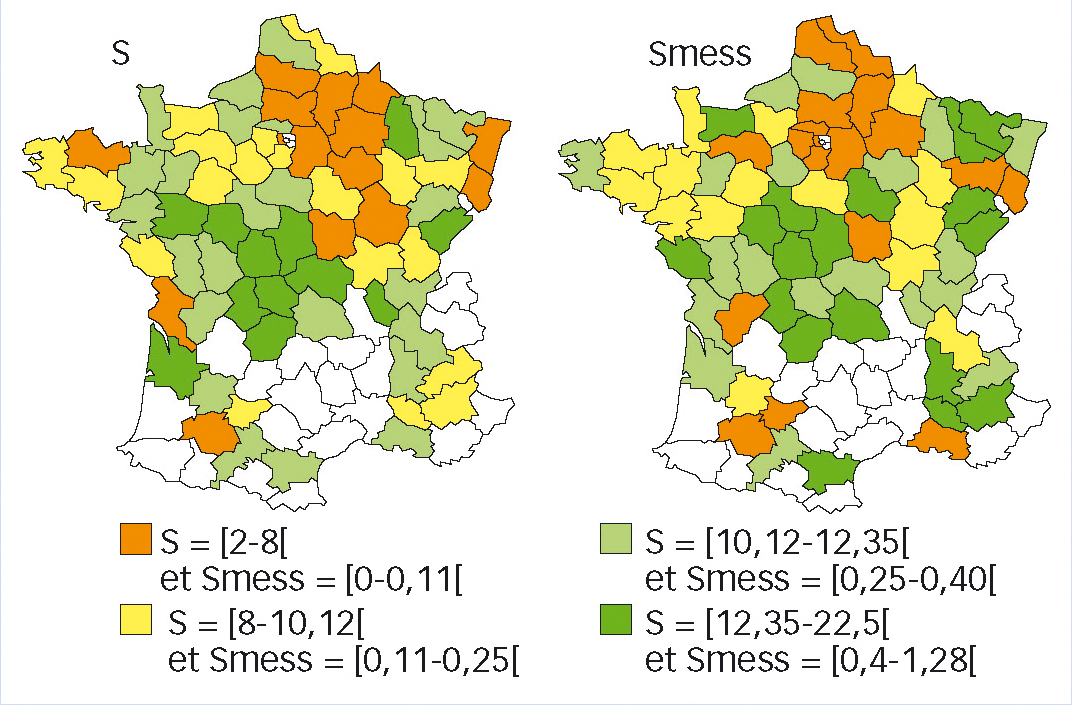
Les couleurs correspondent aux quartiles qui découpent en quatre parties les variables, des valeurs les plus faibles (orange) aux plus élevées (vert foncé).
S. La richesse spécifique est maximale dans le Massif central, le Limousin et le Centre-Ouest, du Cher au Maine-et-Loire. Ces régions contrastent avec un large quart nord-est incluant Île-de-France, Picardie, Champagne-Ardenne et Alsace où la flore des parcelles de blé est très pauvre.
Smess. La richesse en espèces messicoles semble mieux faire ressortir l'influence des facteurs édapho-climatiques (beaucoup sont favorisées en situation xéro-thermophile sur sols calcaires superficiels), avec davantage de messicoles sur les couches géologiques du Jurassique (de la Charente-Maritime au plateau lorrain) ainsi qu'en moyenne montagne méditerranéenne (Vaucluse, Alpes-de-Haute-Provence /Lubéron).
Fig. 2 : Influence de la région et du traitement
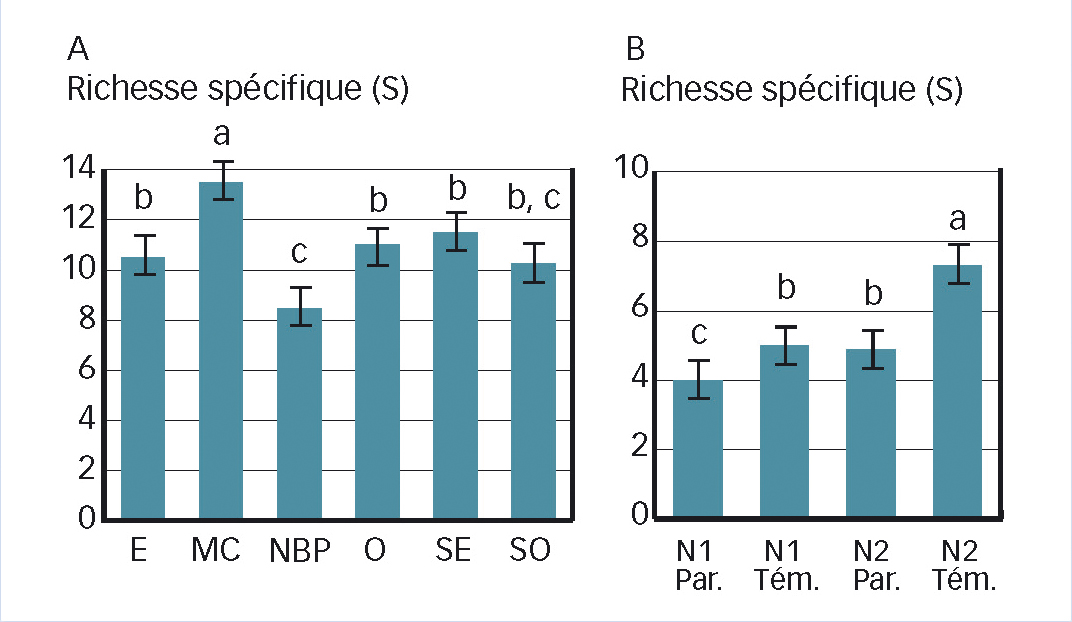
Richesse spécifique moyenne par parcelles : (A) à l'échelle de la région ; (B) par relevés en fonction du traitement : (Par. = traité. Tém. = témoin) et de la saison (N1 automne, N2 printemps). Les lettres a, b et c indiquent des différences significatives
Fig. 3 : Sept groupes d'espèces différenciés
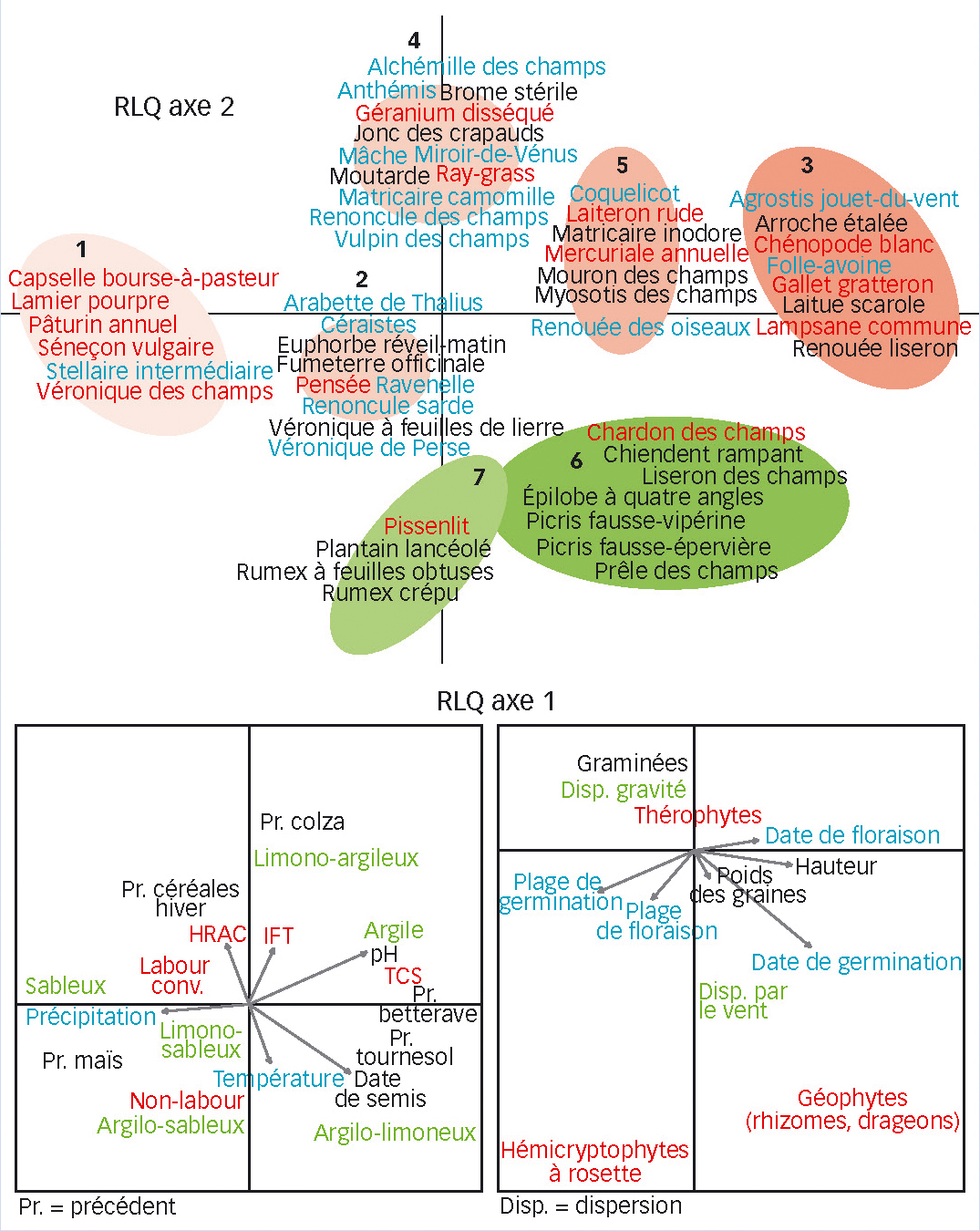
Représentation graphique simplifiée de l'analyse RLQ avec les sept groupes fonctionnels regroupant les espèces en fonction de leurs traits et de leur réponse aux pratiques agricoles et au milieu. En bleu, les espèces en régression, en noir, les espèces stables et en rouge celles en progression, entre 1973 et 2008.
Le Point de vue de
POUR EN SAVOIR PLUS
AUTEURS : *G. FRIED, Anses, Laboratoire de la santé des végétaux, unité Entomologie et plantes invasives, 34988 Montferrier-sur-Lez.
**B. CHAUVEL, **S. GABA, **X. REBOUD, Inra, UMR 1347 Agroécologie, 21065 Dijon.
***J. JULLIEN, DGAL/SDQPV, Draaf/Sral des Pays de la Loire, 49044 Angers.
****P. POINTEREAU, Solagro, 75, Voie du Toec, 31000 Toulouse.
*** **A. RODRIGUEZ, Acta, 31450 Baziège.
CONTACT : guillaume.fried@anses.fr
BIBLIOGRAPHIE :La bibliographie de cet article (9 références) est disponible auprès de son premier auteur (contact ci-dessus).

