 Imprimer
Imprimer
Plants de tomate sous serre. La recherche d'auxiliaires utilisables contre Tuta absoluta passe par des tests sur plants, indispensables après les travaux en laboratoire. Photo : É. Tabone
La tomate est cultivée dans le monde sur plus de 4,5 millions d'hectares (Tropea Garzia et al., 2012), avec 145 millions de tonnes de tomates produites par an dont 90 % dans l'hémisphère Nord (Faostat, 2011). Cette culture de grande importance économique est menacée par de nombreuses maladies et ravageurs – dont Tuta absoluta...
Pourquoi tester des trichogrammes
Tuta absoluta, une mineuse très nuisible
Le lépidoptère Tuta absoluta (Meyrick 1917) (Lepidoptera : Gelechiidae) est un ravageur envahissant originaire d'Amérique du Sud qui a colonisé rapidement le Bassin méditerranéen (Urbaneja et al., 2007 ; Desneux et al., 2010 ; Lacordaire & Feuvrier, 2010 ; Tropea Garzia et al., 2012).
Les adultes pondent leurs œufs sur les plants. Les larves creusent des galeries ou mines dans le parenchyme de tous les organes aériens de la plante. Les dégâts sont directs si la larve s'attaque au fruit ou indirects via la réduction du rendement photosynthétique des feuilles attaquées (Pereyra & Sánchez, 2006 ; Germain et al., 2009). Les pertes peuvent aller jusqu'à 100 % en l'absence de traitement(Gonzales-Cabrera et al., 2011).
Se tourner vers le biocontrôle
Des stratégies de protection doivent être développées pour maîtriser les populations. Mais le comportement « mineur » des chenilles rend difficile le contrôle de ce ravageur (Frandon et al., 2010 ; Polaszek et al., 2012).
Des produits phytopharmaceutiques sont autorisés et utilisés, mais certains présentent des risques tant pour la santé des agriculteurs et des consommateurs que pour la biodiversité (Branco & Franca, 1993). De plus, T. absoluta a déjà développé des résistances à certains insecticides (Lietti et al., 2005). Enfin, dans le cadre du plan Ecophyto, la réduction d'usage des pesticides est recherchée (Frandon et al., 2010(1)).
Des méthodes de lutte biologique et de protection intégrée contre T. absoluta existent mais elles doivent être développées pour plus d'efficacité. La protection biologique et la protection intégrée, basées sur des méthodes préventives et curatives (Tabone et al., 2012), sont utilisées sur tomate depuis les années 1980 (Trottin et al., 2012). Citons les variétés résistantes, la prophylaxie (filets anti-insectes aux entrées, destruction des plants infestés) et le suivi des populations grâce à des pièges à phéromones (Germain et al., 2012 ; Frandon et al., 2010). Il existe aussi des traitements à base de Bacillus thuringiensis, de nématodes ou d'extraits de plantes, ainsi que le piégeage de masse et l'usage d'auxiliaires prédateurs ou parasitoïdes (González-Cabrera et al., 2011 ; Batalla-Carrera et al., 2010 ; Ferreira et al., 2012 ; Desneux et al., 2010 ; Urbaneja et al., 2009 et Séguret et al., 2011 ; respectivement).
L'intérêt des parasitoïdes
Les insectes parasitoïdes effectuent leur développement dans un insecte-hôte dont ils se nourrissent, entraînant sa mort. Ce sont de bons auxiliaires (Badendreier et al., 2003).
Les femelles parasitoïdes oophages pondent leurs œufs dans ou sur les œufs de leur hôte. Ainsi, contre T. absoluta, ce type de parasitoïde va entraîner la mort du ravageur avant l'apparition du stade larvaire responsable des dégâts (Frandon et al., 2010 ; Do Thi Khanh et al., 2012). Les trichogrammes Trichogramma spp. sont des parasitoïdes oophages, et certaines souches au sein de ces espèces parasitent naturellement les œufs de T. absoluta (Polaszek et al., 2012).
Pourquoi chercher de nouvelles espèces de parasites oophages ?
L'efficacité des trichogrammes contre T. absoluta a été montrée en Amérique du Sud où Trichogramma pretiosum est utilisé en culture de tomate (Faria et al., 2008).
En Europe, Trichogramma achaeae est commercialisé après de bons résultats comme auxiliaire contre T. absoluta en laboratoire et en champ (Cabello et al., 2009). Mais ses difficultés d'élevage et de stockage augmentent les coûts de production. En effet, l'espèce n'a pas de capacité de diapause, ce qui empêche son stockage au froid (Do Thi Khanh et al., 2012). De plus, jusqu'en 2014, l'évolution réglementaire (liée au décret 2012-140) faisait craindre que la commercialisation de cette espèce exotique soit bloquée.
D'où le programme TutaPI
Afin de développer les moyens biologiques contre T. absoluta, le ministère chargé de l'Agriculture a financé le programme Casdar (compte d'affectation spécial pour le développement agricole rural) TutaPI (2011-2014) : « Recherche et intégration d'une protection biologique contre Tuta absoluta, ravageur invasif de la tomate ». Ce projet a associé des partenaires de la recherche, de l'expérimentation, du développement et des producteurs. L'un de ses axes consistait à chercher des espèces de trichogrammes indigènes plus efficaces et/ou plus économiques que la référence T. achaeae déjà commercialisée.
Le travail réalisé
Collecte du matériel biologique
Des trichogrammes ont été récoltés dans différentes régions françaises, dans des cultures de tomate conduites en agriculture biologique chez des agriculteurs partenaires n'ayant pas réalisé de lâcher de trichogrammes dans leurs cultures.
Pour ce faire, des œufs d'Ephestia kuehniella, collés sur bandelettes de carton, sont placés durant 48 heures sur des plants de tomate en plein champ, sous abris ou serre. Les bandelettes sont ensuite placées à 25 °C jusqu'à l'émergence des trichogrammes. Ces derniers sont identifiés (niveaux morphologique et moléculaire) et séparés par espèce. Des souches de plusieurs espèces sont ainsi récoltées.
Parallèlement, d'autres souches sont choisies parmi la collection vivante de trichogrammes maintenue en permanence au sein du laboratoire biocontrôle de l'Inra UEFM.
Élevage et tests à l'Inra UEFM
Les souches de trichogrammes collectées sont élevées en tubes contenant du miel, sur des œufs d'E. kuehniella irradiés (pour empêcher l'éclosion de leurs larves) collés sur une bandelette de carton, dans une pièce à 25 °C (±1 °C) de température, 60 % (±10 %) d'humidité relative avec une photopériode lumière 16 heures/obscurité 8 heures.
Nous avons testé en laboratoire le parasitisme exercé sur les œufs de T. absoluta par les souches élevées.
Les neuf souches les plus performantes à l'issue de ces tests, appartenant à cinq espèces différentes, ont été sélectionnées pour des tests plus proches des conditions de culture.
Tests en serres expérimentales
Ces tests ont été réalisés dans des serres expérimentales sur culture de tomate. Le taux de parasitisme des souches sélectionnées a été comparé à celui de la souche de référence de T. achaeae testée dans les mêmes conditions.
Comme ces neuf souches présentent des facilités d'élevage et de stokage permettant de réduire les coûts de production, celles au taux de parasitisme supérieur ou égal à celui de T. achaeae pourront constituer des alternatives économiquement intéressantes (Do Thi Khanh et al., 2012).
Repérage de conditions d'efficacité
Par ailleurs, certaines conditions environnementales et différents facteurs pouvant influencer le comportement des trichogrammes ont été relevés, et leur effet sur le taux de parasitisme des trichogrammes testé.
En effet, la connaissance de ces effets sur la performance des souches peut permettre d'optimiser les méthodes de lâchers et les modalités d'utilisation des auxiliaires en fonction de la souche lâchée et des conditions de l'environnement.
Cette étude est la dernière étape avant des essais, en conditions de cultures et en serres de production, des souches de trichogrammes présentant le meilleur potentiel comme auxiliaire pour la protection biologique contre T. absoluta.
Protocole expérimental
Plusieurs essais ont été réalisés en 2012. À chaque essai, la souche commercialisée T. achaeae « CQ » a servi de référence. Dix séries ont été réalisées d'avril à octobre. En tout, dix souches (celle de T. achaeae et neuf autres) ont été testées.
Organisation des compartiments
Chaque souche a été étudiée dans un compartiment sous serre avec six lignes de culture (dont deux de bordure), chacune de quatorze jeunes plants de tomates (âgés de quatre semaines) de variété Climberley (Syngenta).
Quinze des 84 plantes de chaque compartiment étaient infestées artificiellement par T. absoluta. Les autres plants miment la réalité d'une culture de tomates en serre (Figure 1 page suivante).
Ponte de T. absoluta
Les plants à infester ont été mis en présence d'adultes de T. absoluta dans des cages durant quelques heures (souches réparties au hasard, durée d'infestation ajustée au nombre d'adultes par cage).
Puis les plants ont été retirés des cages en veillant à ce qu'aucun ravageur ne reste sur eux. La présence d'au moins deux œufs par plant a été vérifiée avant leur mise en place dans les compartiments.
Lâchers des trichogrammes
Les trichogrammes ont été fournis par l'Inra UEFM sous forme d'œufs d'E. kuehniella parasités par une souche, collés sur une bandelette de carton sans miel pour reproduire les conditions de lâchers du commerce. Environ 2 500 œufs parasités sont présents sur chaque bandelette, soit le double de la dose recommandée car le nombre d'œufs de T. absoluta par compartiment (jusqu'à près de 400 œufs pour 50 m² de culture) était bien supérieur à une situation moyenne en production.
Dès l'apparition des premières émergences de trichogrammes, la bandelette de carton avec les œufs parasités a été placée au centre du compartiment, à l'abri du soleil pour éviter le dessèchement des œufs et la mort prématurée des parasitoïdes. Les trichogrammes ont été laissés durant sept jours. Les températures moyennes du jour et de la nuit ont été relevées ainsi que l'humidité relative moyenne.
Pour les quatre premières séries d'essais, les mêmes plantes non infestées ont été utilisées pour deux séries consécutives (séries 1 et 2 d'une part, 3 et 4 d'autre part).
Vu la croissance des plants de tomates, ceux-ci étaient donc plus grands lors des séries 2 et 4. À partir de la série 5, les plants non infestés ont été changés à chaque série afin que leur taille soit comparable entre les séries.
Incubation et comptage
Après sept jours, les quinze plantes avec des œufs de T. absoluta sont retirées des compartiments et stockées à 25 °C. Après cinq jours d'incubation, le nombre d'œufs parasités (noirs), non parasités (blancs, desséchés...) ainsi que le nombre de chenilles par feuille et par plante sont déterminés à la loupe frontale et notés.
Un taux de parasitisme est calculé par plante à l'aide de la formule suivante :
Taux de parasitisme = (nombre d'œufs noirs parasités)/(nombre total d'œufs). La qualité des trichogrammes a été vérifiée après chaque transport par le calcul du taux d'émergence.
Analyses statistiques
Les analyses statistiques ont été réalisées avec le logiciel R version 3.0.0 (R Core Team, 2013).
Dans un premier temps, le taux de parasitisme a été mesuré par plant (n = 15 plants avec une à cinq séries pour chaque souche et dix répétitions pour la référence T. achaeae). Ensuite, une moyenne a été effectuée sur l'ensemble des résultats (toutes séries confondues) pour chacune des souches.
Les différences des taux de parasitisme entre souches, entre séries et entre distances au point de lâcher des plants infestés ont été analysées à l'aide d'un modèle linéaire généralisé GLM avec une distribution de Quasi-Poisson (données issues de comptages). La correspondance au modèle a été testée grâce à un test F (model : quasipoisson, link : log).
Les différences significatives entre les taux de parasitisme des différentes souches ont été étudiées à l'aide d'un test de Wilcoxon pour comparaisons multiples. Ces tests ont été réalisés pour les différentes souches à partir des différents paramètres mesurés au cours de l'expérimentation.
Les corrélations du taux de parasitisme ont été étudiées à l'aide du coefficient de corrélation de Spearman (ρ = coefficient de corrélation), avec :
la température moyenne du jour ;
– l'humidité relative ;
– la date de la série ;
– la distance au point de lâcher des trichogrammes ;
– le nombre d'œufs de T. absoluta sur les plants ;
Pour les corrélations significatives, des régressions linéaires ont été recherchées.
Afin d'identifier un potentiel effet des différentes souches testées selon les séries sur ces corrélations, les tests se sont référés à T. achaeae, testée à chaque série. Vu la forte dispersion et la non-normalité fréquente des taux de parasitisme, ils sont représentés par des diagrammes en boîtes (Figure 2).
Les résultats
Efficacité des souches
Il y a un effet significatif de la souche considérée sur le taux de parasitisme (p = 2,2 x 10-16). Les souches X, BU, AZ, BV et CV ont un taux de parasitisme moyen significativement plus faible que celui du témoin T. achaeae CQ (respectivement ; p = 0,0025 ; p = 1,2 x 10-6 ; p = 5,9 x 10-5 ; p = 0,00081 et p = 0,00031). En revanche, les quatre souches CX, DF, CK et CL sont statistiquement équivalentes au témoin CQ (T. achaeae).
Conditions expérimentales : effets de la température, de l'humidité, de la distance au point de lâcher
Dans cet essai, le compartiment dans lequel elle a été testée n'a pas d'effet significatif sur le taux de parasitisme de la souche (p = 0,36). Il en est de même pour la densité d'œufs de T. absoluta sur les plants infestés (ρ = 0,06 ; p = 0,17).
Une corrélation (coefficient ρ) existe entre l'avancement dans les séries et deux données (elles-mêmes très corrélées entre elles) : les températures moyennes du jour et l'humidité relative (ρ = 0,048 ; p = 2,2 x 10-16 et ρ = 0,75 ; p = 2,2 x 10-16 respectivement). Le taux de parasitisme est corrélé positivement avec l'augmentation de la température moyenne du jour et de l'humidité relative (respectivement ; ρ = 0,21 ; p = 2,3 x 10--7 et ρ = 0,19 ; p=2,3 x 10-7). Les corrélations entre le taux de parasitisme de T. achaeae et, d'une part le numéro de la série, d'autre part l'humidité relative, sont significatives (ρ = 0,33 ; ρ = 0,36 et p = 2,7 x 10-5 ; p = 5,7 x 10-6 ; respectivement). En revanche, le taux de parasitisme de T. achaeae n'a pas de corrélation significative avec la température moyenne (ρ = 0,08 ; p = 0,32).
Enfin, le taux de parasitisme de l'ensemble des souches décroît avec l'augmentation de la distance du plant de tomate infesté au point de lâcher (ρ = -0,20 ; p = 8,05 x 10-7).
Lorsque les souches sont considérées sans la référence T. achaeae, cette corrélation négative est également significative (ρ = -0,23 et p = 5,0 x 10-7).
En revanche pour T. achaeae, le taux de parasitisme sur le plant infesté n'est pas significativement corrélé avec la distance au point de lâcher (ρ = -0,12 ; p = 0,13) : il n'existe aucune différence significative entre les taux de parasitisme de T. achaeae observés en fonction des plants étudiés.
Quelles conclusions en tirer ?
Quatre souches à suivre
Parmi les souches testées, trois souches autochtones parmi les quatre ayant un taux de parasitisme équivalent à T. achaeae (CQ) pourront être particulièrement étudiées pour voir si elles peuvent constituer des alternatives pour le contrôle de T. absoluta. Toutes peuvent être élevées facilement sur des œufs d'E. kuehniella (taux de parasitisme élevé sur cet hôte, adultes émergents de bonne qualité) avec possibilité de ralentissement de développement (diapause pouvant permettre le stockage au froid), donc un coût de production inférieur à T. achaeae.
Conditions environnementales : un effet classique de la température
La corrélation positive entre le taux de parasitisme des trichogrammes et la température moyenne du jour indique que, comme la plupart des insectes, leur activité augmente avec l'élévation de la température. De fait, selon Suverkropp et al., (2010), le temps de recherche des œufs (d'Ostrinia nubilalis) par les trichogrammes est environ deux fois plus court à 25 °C qu'à 18 °C.
L'humidité et la lumière jouent aussi
Il existe aussi une corrélation positive entre le taux de parasitisme des trichogrammes des différentes souches et l'humidité relative dans le compartiment. Des valeurs entre 60 et 80 % entraînent un taux d'émergence supérieur des trichogrammes lâchés, et ils sont de meilleure qualité avec une fécondité augmentée (Calvin et al., 1984 ; Gross, 1988).
Ainsi, la corrélation positive entre le taux de parasitisme et l'avancement dans les séries au cours du temps est en partie expliquée par l'augmentation des températures moyennes du jour et de l'humidité relative (Suverkopp et al., 2010). Ceci est confirmé lors des séries 2 et 3 pour lesquelles il y a une baisse significative du taux de parasitisme parallèle à une forte chute des températures moyennes du jour.
Au cours de l'avancement dans la saison, la photopériode et l'intensité lumineuse ont augmenté. Les trichogrammes ayant un comportement strictement diurne, l'augmentation de la photopériode leur donne plus de temps pour parasiter les œufs du ravageur. L'augmentation de l'intensité lumineuse a aussi un effet positif sur l'activité et le taux de parasitisme des trichogrammes (Costas, 1951).
La taille des plants n'aurait pas d'effet significatif
Dans nos conditions expérimentales, il ne semble pas y avoir d'effet de la taille de ces plants sur la performance des trichogrammes. En effet, il n'y a pas de différence de taux de parasitisme entre les séries 1 et 4 où les plants sains sont petits et grands respectivement. De même, les taux de parasitisme des séries 2 et 3 sont comparables malgré les tailles différentes de ces plants.
En revanche, il y a une différence significative entre les taux de parasitisme observés entre d'une part les séries 1 et 4 et d'autre part les séries 2 et 3. Ainsi, bien que plusieurs études montrent que l'augmentation de la hauteur et de la surface foliaire entraîne une diminution du parasitisme (Ables et al., 1980 ; Wang et al., 1997), les conditions climatiques ont certainement eu une influence bien plus forte sur l'activité des trichogrammes que la structure des plants.
Effet de la distance au point de lâcher : les souches indigènes se dispersent moins que T. achaeae
Le taux de parasitisme des souches de trichogrammes testées diminue avec la distance au point de lâcher ; il est corrélé négativement avec la distance au point d'émergence des adultes. En revanche, dans cette expérience, le taux de parasitisme de T. achaeae seul ne diminue pas significativement en s'éloignant du point de lâcher. La capacité de dispersion moyenne de cette espèce semble supérieure à 4 mètres (distance maximale d'un plant infesté au point de lâcher).
Pour les autres souches testées en serre, il semble que la distance de dispersion moyenne soit inférieure. Pour ces autres souches, des différences significatives entre les taux de parasitisme des plants infestés à différentes distances n'ont été mises en évidence qu'entre des plants parmi les plus proches du point de lâcher (plants n° 6 et 9, Figure 1) et les plants les plus éloignés et situés du côté du compartiment où se trouve le système de ventilation (plants n° 2 et 15) (Figure 1). C'est donc l'association de distances importantes au point de lâcher (3,9 et 3,4 mètres) et d'un déplacement à contre vent qui limite l'accès des trichogrammes à ces plants et réduit leur taux de parasitisme.
Perspectives pour la protection biologique contre T. absoluta
Souches autochtones : inutile d'augmenter la dose si le nombre de points de lâcher reste le même
Les expériences en serre ont montré que plusieurs souches autochtones ont une efficacité comparable à celle de T. achaeae. Ce dernier étant environ 1,3 fois plus cher à produire que les autres espèces (Do Thi Khanh et al., 2012), il est donc envisageable de lâcher, pour un même coût, 30 % de plus d'individus des souches autochtones si les résultats sont comparables voire supérieurs à ceux fournis par T. achaeae. Deux séries d'essai ont été réalisées en 2013 en serres expérimentales sur le centre Ctifl de Balandran, selon un protocole similaire à celui présenté ici, avec une des souches les plus efficaces introduite à une dose supérieure de 30 %. L'augmentation de dose n'a pas augmenté significativement le taux moyen de parasitisme dans cet essai (Trottin, données non publiées).
Ces résultats et l'absence d'effet de la distance au point de lâcher sur le parasitisme de T. achaeae suggèrent que l'efficacité supérieure de T. achaeae pourrait provenir de sa meilleure dispersion (Tabone et al., 2012).
Points à évaluer demain
Lors de prochaines expériences en serre, il serait intéressant de récupérer des données telles que le pattern de ponte de T. absoluta sur les plants de tomates et celui de l'oviposition des trichogrammes des différentes souches. Cela permettrait de mieux comprendre le comportement du ravageur et celui des auxiliaires et d'optimiser les méthodes de lâcher de ces derniers.
En effet, le pattern de ponte de l'hôte est primordial dans la préférence et l'efficacité des trichogrammes (Wajnberg et al., 2003 ; Faria et al., 2008 ; Chailleux et al., 2012).
De plus la structure de la plante-hôte (ex. : présence et densité des trichomes ou « poils » des plants de tomate) influence le comportement de recherche et les déplacements des parasitoïdes donc leur taux de parasitisme (Kashyap et al., 1991 ; Romeis et al., 1997 et 1998 ; Faria et al., 2008 ; Tabone 2010). Faria et al. (2008) ont observé que l'apex des plants de tomate est une zone refuge pour les œufs du ravageur face au parasitisme de T. pretiosum. Les œufs de T. absoluta pondus à cet endroit ne sont pas parasités par les femelles de T. pretiosum : la plus forte densité de trichomes ralentit les déplacements des trichogrammes et peut les piéger par leur exsudat.
Conclusion
Dans le cadre du projet TutaPI, ces essais en serres expérimentales font suite aux expériences en tubes et en mésocosmes effectuées par le laboratoire Biocontrôle de l'Inra UEFM. Deux étapes préliminaires avaient permis de choisir neuf nouvelles souches (d'espèces autres que T. achaeae) avec le meilleur potentiel contre T. absoluta.
Les expériences en serre expérimentales ont permis de tester la performance de ces souches dans des conditions plus proches de celles de la production. Trois souches autochtones ayant un taux non significativement différent de T. achaeae pourraient représenter une alternative intéressante.
Plusieurs conditions expérimentales ont influencé le comportement et la performance des trichogrammes. La distance entre le point de lâcher des auxiliaires et les plants infestés (comprise entre 0,5 et 3,95 mètres) est corrélée négativement avec le taux de parasitisme des nouvelles souches, alors qu'elle n'influence pas le taux de parasitisme de T. achaeae. La performance de ce dernier pourrait avoir comme origine ses capacités de dispersion supérieures. Les souches autochtones, avec un taux de parasitisme comparable à celui de T. achaeae, semblent avoir une dispersion plus faible.
Des stratégies de lâchers adaptées (points plus rapprochés) pourraient-elles en faire des alternatives au moins aussi efficaces que T. achaeae ? Si ces souches ont un coût de production inférieur, cela compenserait l'augmentation des coûts de lâchers. Afin de compléter ces observations, une étude de la dispersion des souches en laboratoire devra être envisagée.
<p>(1) http://agriculture.gouv.fr/ecophyto</p>
Fig. 1 : Dispositif expérimental
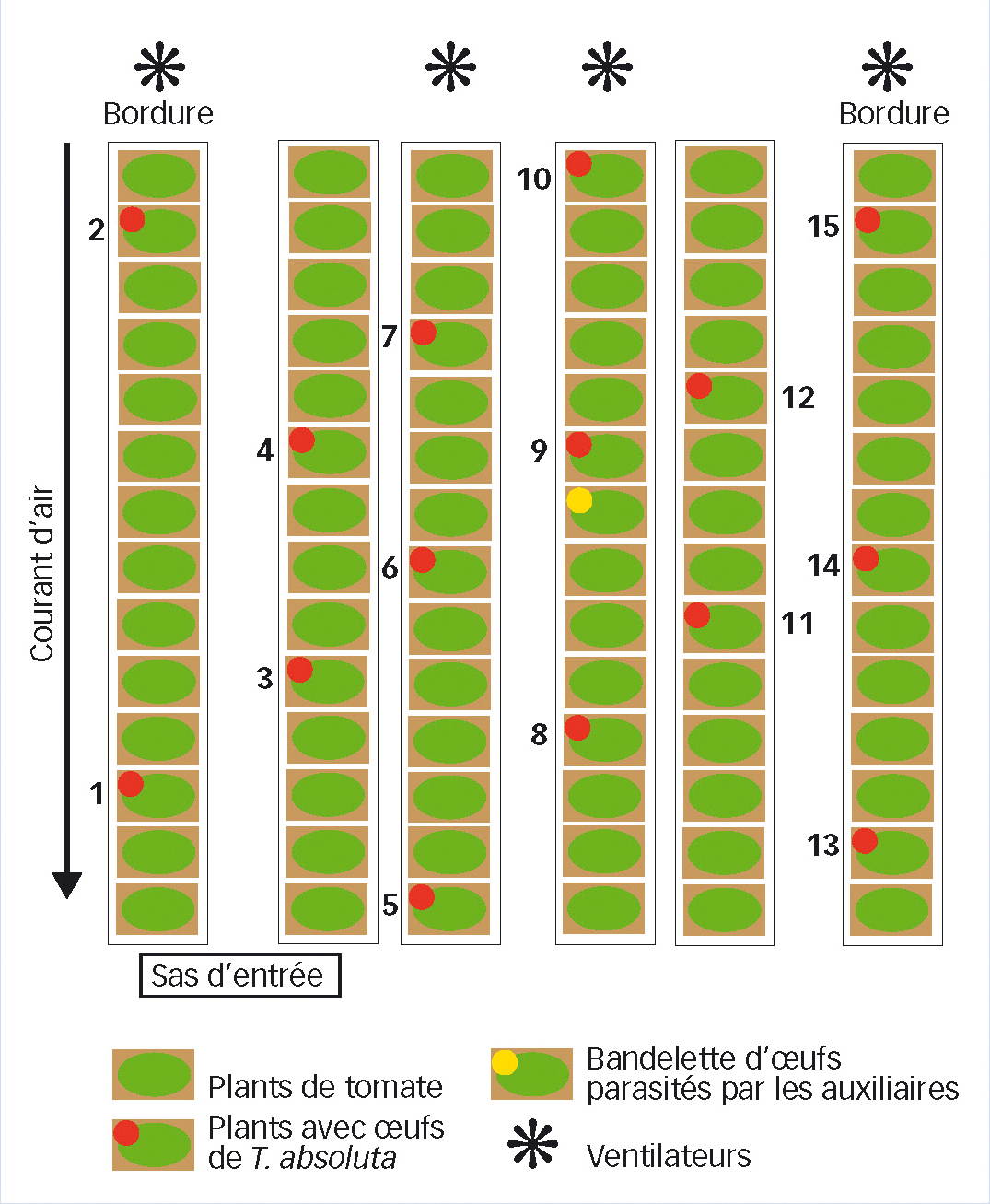
Plan d'un compartiment. Les quinze plants, numérotés de 1 à 15 pour chaque série, sont ceux préalablement infestés par Tuta absoluta. Chacune des dix souches (T. achaeae et les neuf souches sous numéro) a été testée dans quatre compartiments.
Fig. 2 : Taux de parasitisme des différentes souches de trichogrammes obtenus lors des expériences en serre (classés par ordre croissant)
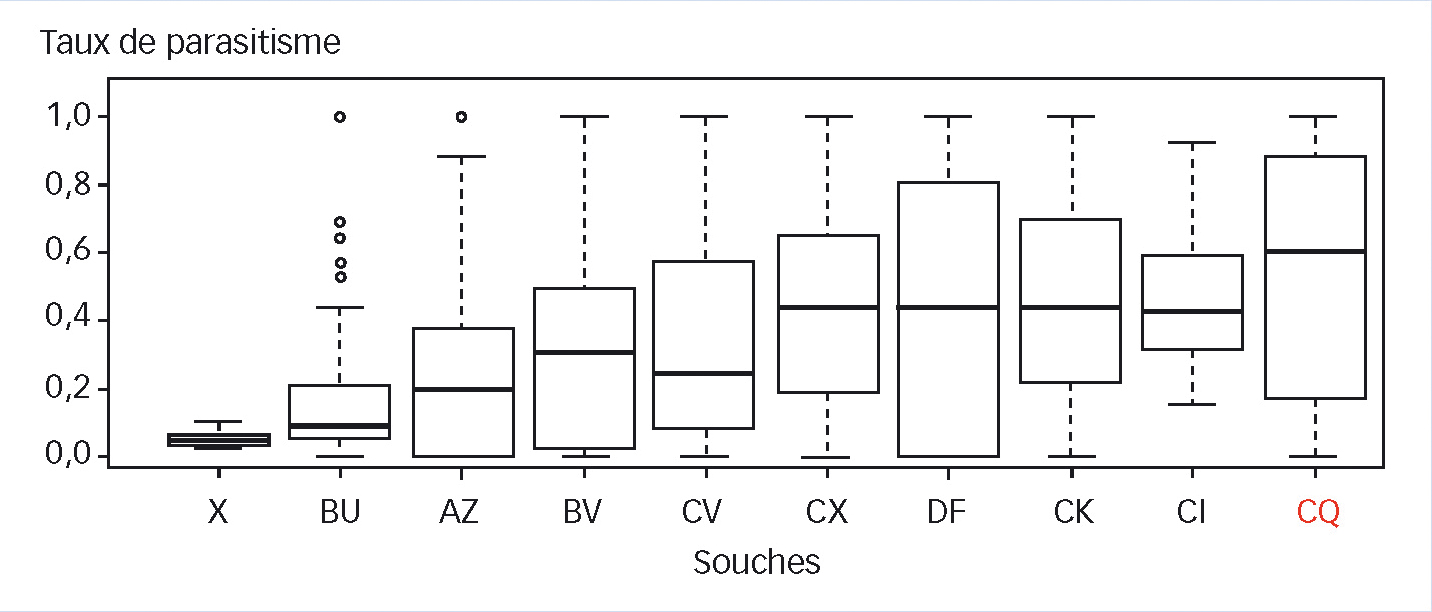
Pour chaque souche, le taux de parasitisme correspond à la moyenne des résultats par plant, toutes séries confondues. Les souches X, BU, AZ, BV et CV sont statistiquement différentes de CQ (= T. achaeae), au contraire des souches CX, DF, CK et CL.

