 Imprimer
Imprimer
Symptômes de rouille jaune dus à la race Warrior : les stries jaunes d'urédospores « classiques » s'accompagnent de téleutosores brunes, typiques de Warrior. Photo : Inra
L'importance de l'épidémie de rouille jaune du blé en 2014, nettement supérieure à celle observée en 2013, a surpris et inquiété la profession agricole. Les causes de cette flambée sont intéressantes à connaître. Nous verrons qu'elle résulte de la combinaison de trois facteurs :
1/ la climatologie, la température hivernale ayant favorisé une attaque très précoce ;
2/ la culture de variétés sensibles ;
3/ la présence d'inoculum virulent.
De plus, les résultats des essais variétaux, menés dans le cadre du FSOV Rouille Jaune, suggèrent l'existence d'une régionalisation des pressions de maladie voire des races du pathogène présentes en France.
Rappel utile sur le parasite responsable de la rouille jaune du blé
Une succession rapide de cycles infectieux
Le champignon responsable de la rouille jaune du blé, maladie dévastatrice mondialement, est un basidiomycète biotrophe, Puccinia striiformis f. sp. tritici (Pst). Ce pathogène ne se développe que sur des tissus vivants de la partie aérienne de la plante. Il infecte les feuilles quand la température est fraîche et l'hygrométrie élevée.
La maladie est caractérisée par sa propagation en foyers et sa grande sensibilité aux conditions climatiques qui la rend sporadique, son intensité variant d'une saison à l'autre.
Les épidémies sont déclenchées par des urédospores, spores asexuées dicaryotiques (ayant deux noyaux haploïdes). Le cycle infectieux (succès de l'infection des urédospores, période de latence entre la pénétration des spores et la sporulation, taux de sporulation) est influencé par les conditions climatiques et la sensibilité de la variété.
L'épidémie est définie par la succession de cycles infectieux, chacun durant environ deux semaines en conditions favorables. Pst a un développement rapide à l'intérieur des tissus-hôtes par des lésions qui croissent entre les nervures de la feuille et produisent de grandes quantités de spores. Les stries sporulantes sont caractéristiques de la rouille jaune, à la différence de la rouille brune aux lésions rondes et de taille limitée.
Dispersion de la maladie par les spores
Les spores sont dispersées par le vent sur de courtes ou de très longues distances.
Un mucilage entourant les urédospores permet dans des conditions humides de grouper deux à neuf spores en un amas limitant ainsi la distance de dispersion. Plus légères, les spores isolées produites dans des conditions plus sèches sont plus aptes à la dispersion à longue distance.
Les années de sévères épidémies combinent généralement la présence d'un inoculum primaire important et des températures hivernales supérieures à la normale permettant le démarrage précoce de l'épidémie. Les conditions favorables du printemps généralisent l'épidémie, apparue initialement en foyers.
La phase sexuée de Pst sur un hôte alternant, l'épine vinette (Berberis spp.), a été décrite récemment en conditions de laboratoire mais n'a pas été observée dans la nature. L'espèce a été éradiquée en France pour lutter contre la rouille noire, et la réglementation impose que les Berberis ornementaux soient résistants à la rouille. Il est déterminant de maintenir cette réglementation.
Les souches de Pst évoluent dans le temps
Lignées « nordique » et « méditerranéenne »
Pst sélectionne des virulences capables de contourner les gènes de résistance spécifique portés par les variétés de blé sélectionnées pour leur résistance. Les races de Pst responsables des épidémies en France appartiennent à deux lignées clonales. L'une, typique du nord-ouest de l'Europe, est caractérisée par un grand nombre de virulences et adaptée aux températures fraîches. L'autre, typique du Bassin méditerranéen, a peu de virulences et est adaptée à plus haute température.
La souche PstS1/S2 n'est pas présente en France
Depuis 2000, une souche agressive et tolérante à haute température (PstS1/S2) a émergé dans de nombreux pays de plusieurs continents (Hovmøller et al., 2011). Toutefois, PstS1/S2 ne possède pas les virulences Vr1, Vr3, Vr17 et Vr32 nécessaires pour contourner la résistance de la majorité des variétés françaises et ne s'est pas établie sur notre territoire (de Vallavieille-Pope et al., 2012).
Warrior attaque depuis 2011
Une autre race invasive d'origine exotique, la race Warrior, a été détectée simultanément dans les pays de l'Europe de l'ouest en 2011. Elle possède les virulences 7 et Sp, plus celles portées par la race Oakley prédominante jusqu'en 2011.
Depuis lors, Warrior domine la population française de Pst (www.wheatrust.org). Les observateurs décrivent des symptômes différents des autres races : peu de sporulation, présence de chloroses et nécroses, formation abondante de téleutosores.
Blé et gènes de résistance
Des gènes de résistance majeurs contournés
L'utilisation de variétés résistantes est le moyen de lutte le plus efficace, économique et respectueux de l'environnement. Certaines variétés sont résistantes dès le stade plantule alors que d'autres acquièrent une résistance au stade adulte.
Soixante-dix gènes de résistance Yr ont été décrits. En France, dix principaux gènes de résistance majeurs Yr exprimés au stade plantule ont été utilisés successivement ou de façon combinée dans les variétés cultivées. L'ensemble de ces gènes ont été contournés (de Vallavieille-Pope et al., 2012) mais d'autres gènes ou combinaisons de gènes demeurent efficaces.
Des résistances tout de même durables... pourquoi ?
Pourtant, la majorité des variétés conservent une résistance adulte importante. Son déterminisme génétique demeure le plus souvent inconnu.
Certaines combinaisons de gènes de résistance ont montré une résistance durable, c'est-à-dire efficace sur de grandes surfaces pendant plus de dix ans sous des conditions épidémiques favorables. C'est le cas de Camp Rémy, Renan, Soissons et Apache qui combinent des gènes de résistance majeurs exprimés dès le stade plantule et des gènes de résistance partielle exprimés au stade adulte (Sørensen et al., 2014).
La résistance d'Apache, efficace depuis plus de quinze ans, résulte du cumul de deux gènes majeurs exprimés dès le stade plantule (Yr7 et Yr17) et d'un QTL (QYr.inra 4B) exprimé au stade adulte (Paillard et al., 2012). L'association reste efficace vis-à-vis de la race Warrior (test au stade adulte, Inra 2012) mais est contournée par la race KWS Sterling présente en Angleterre depuis quatre ans.
Dans le cas d'attaque précoce, des variétés possédant une résistance adulte peuvent montrer des symptômes en début d'épidémie du fait de leur sensibilité au stade jeune.
Quarante-cinq génotypes évalués sur treize sites
Six coopératives et sept obtenteurs
Depuis deux ans, des évaluations variétales vis-à-vis de la rouille jaune en contaminations naturelles sont mises en place par six coopératives des régions traditionnelles de la rouille jaune (départements 14, 22, 59, 76, 80 et 91, voir Figure 1).
Deux types de génotype sont évalués : d'une part des génotypes « révélateurs » choisis car nous connaissons les allèles des gènes majeurs de résistance qu'ils portent ; d'autre part des « variétés récentes » permettant de juger le niveau de résistance dans le matériel français. Au total, 45 génotypes sont évalués.
En parallèle, sept obtenteurs évaluent ces mêmes variétés en inoculant artificiellement la race Warrior (départements 28, 35, 59, 60 et 78).
Malgré des conditions climatiques variées, le témoin sensible Victo, présent sur la quasi-totalité des sites, a subi une intensité de maladie d'au moins 90 % les deux années. L'intensité de maladie était donc suffisante pour tester les résistances.
Mode de classement des variétés
Les notations tardives de l'intensité de maladie (pourcentage de surface foliaire malade) sur les deux premiers étages foliaires (F1 et F2) montrent d'importantes corrélations (r²2013 = 0,87 ; r²2014 = 0,92). Ainsi, l'analyse repose sur la notation d'intensité globale de maladie. Une variété est classée :
- résistante si l'intensité de maladie est inférieure à 3 % (absence de symptômes ou présence de traces) ;
- moyennement résistante si l'intensité est entre 3 % et 15 % ;
- moyennement sensible, si l'intensité est entre 15 % et 40 % ;
- sensible si l'intensité est supérieure à 40 %.
Une nouvelle souche apparue en 2014 ?
En 2013, Warrior est généralisée
L'objectif de cette étude est d'évaluer la stabilité de la résistance variétale à la rouille jaune entre les deux années du projet. Les variétés composant le panel des « génotypes révélateurs » permettent de couvrir les principaux gènes majeurs (Yr3, Yr6, Yr7, Yr17, Yr32) ainsi que d'autres sources (QTL) présentes dans des variétés comme Renan. Une première approche des différents comportements entre génotypes permet ainsi de distinguer les combinaisons de virulences présentes sur chaque site sans avoir recours à une analyse de souches.
En 2013, cette approche en conditions naturelles a fait conclure à la généralisation de la race Warrior sur l'ensemble du territoire. Les analyses en laboratoire de l'Inra ont confirmé le fait.
En 2014, une évolution notable
En conditions naturelles, le comportement des génotypes révélateurs vis-à-vis de la rouille jaune a évolué en 2014. Certains de ces génotypes et certaines variétés récentes présentent une intensité de maladie supérieure à 2013 (Figures 2 et 3).
Cela confirme l'augmentation générale des symptômes liée à la pression rouille jaune, plus forte en 2014 qu'en 2013. Mais cette apparition de nouveaux symptômes est-elle imputable, aussi, à l'apparition d'une nouvelle souche contournant les gènes de résistance ainsi que les sources de résistance au stade adulte de certains génotypes révélateurs non contournés jusqu'à présent ? Cette hypothèse peut être suggérée par l'évolution entre les deux années du projet (Figure 3).
Le seul génotype révélateur ne présentant aucun symptôme maladif pour l'ensemble des sites les deux années est Soissons. À l'exception de Renan et d'Apache, le reste des génotypes révélateurs présentent en 2014 une intensité des symptômes supérieure ou égale à 40 % sur au moins un des sites.
D'après l'analyse menée en laboratoire, la majorité des échantillons prélevés sur l'ensemble des sites les deux années possèdent le même spectre de virulence. Il correspond à la race 239E175V17. Cependant, le spectre d'analyse ne permet pas de différencier la race Warrior de la race KWS Sterling, ni les virulences exprimées au stade adulte. En 2013, le site expérimental de la coopérative Triskalia (22) se distinguait par la présence de la race 110E175 (Avr9, Avr17).
Des souches différentes selon les régions
À l'ouest, du nouveau...
L'étude de la répartition des classes de résistance des génotypes révélateurs selon les sites montre d'importantes différences de répartition entre les sites français à contamination naturelle.
En 2014, les sites de Capseine (76), d'InVivo (91) et de Noriap (80) présentent ainsi moins de variétés classées « sensibles » que les sites d'Agrial (14) et de Triskalia (22).
Ces observations illustrent l'importance de l'interaction variété/site. Par exemple, Camp Rémy et Kranich présentent peu ou pas de symptômes sur les sites de Capseine, InVivo et Noriap, et une intensité globale des symptômes dépassant 50 % sur les sites d'Agrial et de Triskalia.
L'Inra a montré en 2012 l'avirulence de races Warrior et Oakley sur Camp Rémy au stade adulte (Sørensen, 2014). Non contourné en 2013, cette observation supporte l'hypothèse d'apparition d'une nouvelle souche sur ces deux lieux.
Il apparaîtrait donc ainsi une distinction de souches entre, d'une part, les sites d'Agrial et de Triskalia et, d'autre part, les sites d'InVivo, de Noriap et de Capseine. Nous pouvons souligner la logique géographique des deux groupes établis : un groupe à l'ouest (Triskalia-22 et Agrial-14) et un groupe plus à l'est (Capseine-76, Noriap-80 et InVivo-91). Observés uniquement en 2014, ces résultats devront être confirmés en 2015.
Des nouvelles variétés résistantes sur l'ensemble des sites
Malgré l'augmentation générale des symptômes en 2014, les variétés inscrites récemment conservent un comportement intéressant vis-à-vis de la rouille jaune. En 2014, près de 20 % de ces variétés récentes sont résistantes au stade adulte sur tous les sites (inoculation naturelle et artificielle) (voir Figure 2, la variété récente type 2). De plus, plusieurs variétés, contournées sur certains sites, présentent peu ou pas de symptômes sur d'autres (voir Figure 2, une des variétés type 1).
Par ailleurs, ces variétés ont été testées au stade plantule vis-à-vis de douze races, dont Warrior. Plus d'un tiers d'entre elles sont résistantes à l'ensemble des races. Malgré des races de pathogènes au spectre de virulence de plus en plus large, l'innovation variétale reste aujourd'hui le levier agronomique le plus efficace.
Quelles conséquences sur le choix variétal ?
La précocité de l'épidémie est en partie générée par la culture de variétés dépourvues de gènes de résistance efficaces au stade plantule bien que présentant une résistance adulte efficace. L'épidémie est amplifiée par quelques variétés sensibles au stade adulte... Au vu du niveau de résistance élevée de plusieurs variétés récemment inscrites, on se doit d'encourager l'utilisation de la résistance variétale comme un critère déterminant dans le choix variétal.
L'intensité des symptômes au stade adulte est également modulée par la pression de l'inoculum, largement influencée par les conditions climatiques. Ainsi, nous caractérisons les années épidémiques en fonction de l'effet de la température sur la durée de la période de latence. Le nombre de générations mensuelles est évalué pour la période août 2000-février 2015.
Analyse de « l'effet année »
Caractérisation des années par le nombre de cycles infectieux calculé en fonction de la température
En se basant sur le modèle de Zadoks (1971), revu par Rapilly (1976), et les données de températures journalières fournies par Météo France, nous avons calculé le nombre théorique de générations mensuelles sur une cinquantaine de lieux, situés dans des régions traditionnelles de la rouille jaune où les essais des coopératives et des obtenteurs ont été implantés.
La Figure 4 représente le nombre de générations mensuelles pour les campagnes 2013, 2014 et 2015, comparé à des valeurs de référence couvrant la période 2000-2013. Le calcul du nombre de générations mensuelles pour la période de référence des treize années déroulées avant l'étude (août 2000 à juillet 2013, comprenant des années à pressions différentes) permet de définir pour chaque mois de cette période le nombre de générations moyen.
De plus, à chaque mois sont associées une valeur maximale et une minimale correspondant respectivement aux nombres maximal et minimal de générations mensuelles ayant eu lieu le mois considéré sur la période de référence.
Pour rappel, à l'inverse de 2013, les conditions climatiques de la campagne 2014 ont été très propices au développement de la maladie en France, avec une apparition précoce et une forte nuisibilité (« 2014 est sans conteste l'année épidémique la plus importante en surface et en intensité de ces quinze dernières années(1) »). Les sorties du modèle confortent ce constat.
En 2013, le climat explique la faible épidémie
Les conditions hivernales sur les stations suivies en 2013 ont été particulièrement fraîches, avec en moyenne 1,6 °C de moins que la normale les trois premiers mois de l'année, ce qui a limité l'inoculum. De plus, le printemps a été l'un des plus défavorables à la rouille jaune depuis 2001. Le nombre moyen de générations par mois est en effet régulièrement proche du minimum de générations mensuelles observées de 2001 à 2013. Avec moins de quatre générations théoriques entre mars et mai, la multiplication du pathogène au printemps 2013 a ainsi été pénalisée d'une génération par rapport à la normale sur cette période. Cette combinaison du faible inoculum et du développement très limité pour l'ensemble de la période mars/mai 2013 explique la faible épidémie observée.
2014, une année exceptionnelle
En revanche, les conditions automnales normales de 2013, puis l'hiver et le printemps 2014 (de janvier à fin mai), marqués par un nombre de générations mensuelles théoriques toujours proches de la valeur maximale, expliquent l'importance de l'épidémie en 2014. Le caractère exceptionnel de l'année 2014, par ses températures moyennes hivernales et printanières, a permis la multiplication élevée du champignon sans discontinuité. De décembre à mai, le nombre de générations théoriques est ainsi estimé à 8,6 en 2014 contre 6,1 en 2013 et 7,1 en moyenne.
Et pour 2015 ?
À titre indicatif, pour la campagne 2014-2015, le nombre de générations mensuelles théoriques à l'automne a été supérieur à la normale (0,35 génération théorique supplémentaire par rapport à la moyenne entre septembre et novembre), proche du maximum observé depuis la campagne 2001.
Mais les conditions hivernales, normales, sont moins propices qu'il y a un an. Avec 0,55 génération théorique, février 2015 est même plus défavorable qu'en moyenne et nettement moins propice que février 2014 (1,14 génération théorique). Quant au mois de mars, il correspond à une année moyenne.
Bien que les conditions hivernales aient été peu favorables au développement de la rouille jaune, l'importance de l'inoculum primaire - liée à la campagne précédente puis à l'automne très favorables à la maladie - suggère une grande vigilance quant au prochain développement de cette rouille. À moins que les conditions printanières n'en décident autrement...
(1) P. du Cheyron et J.-Y. Maufras (Arvalis) dans la Lettre Yvoir du 14 août 2014.
Fig. 1 : Localisation des essais 2013 et 2014
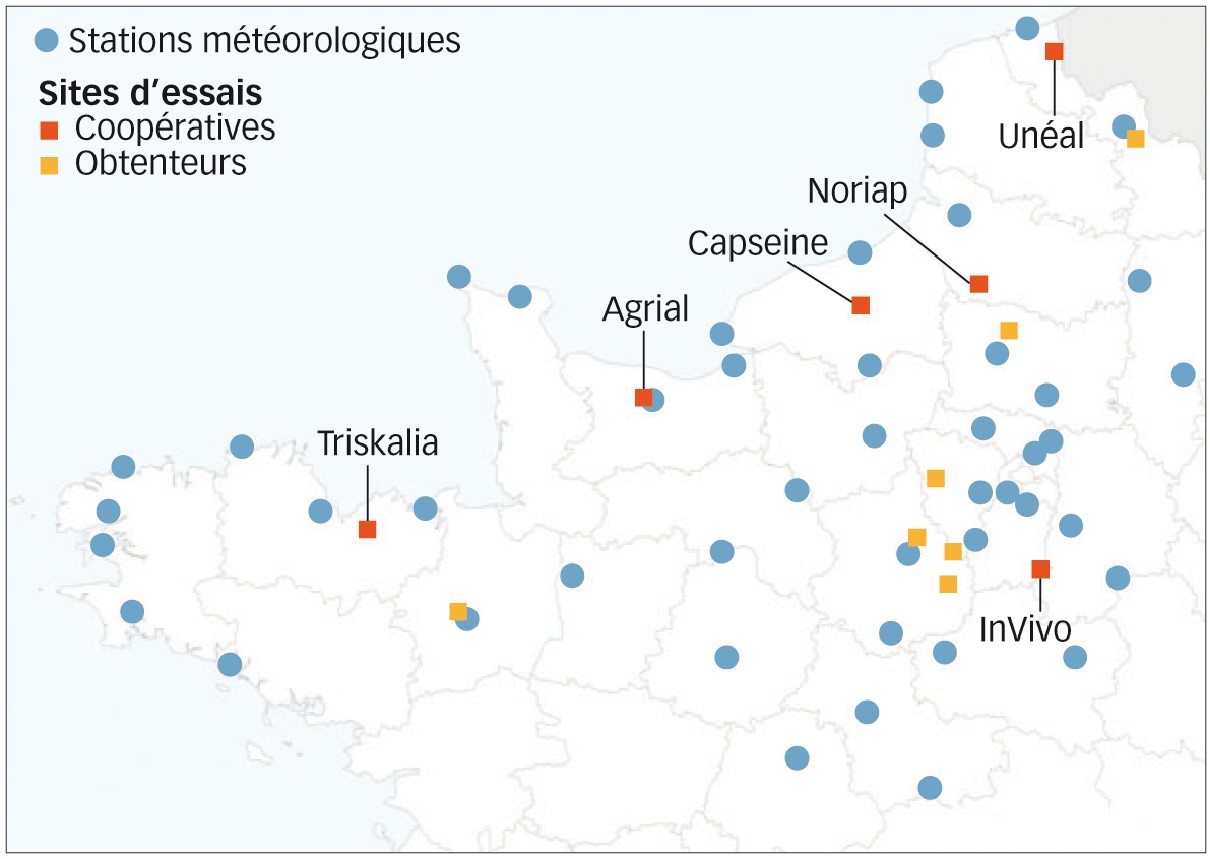
Essais mis en place par les coopératives et les obtenteurs. La carte situe également les stations météorologiques relatives au calcul du nombre théorique de générations mensuelles de Pst.
Fig. 2 et 3 : Stabilité ou instabilité spatiale et temporelle des comportements variétaux
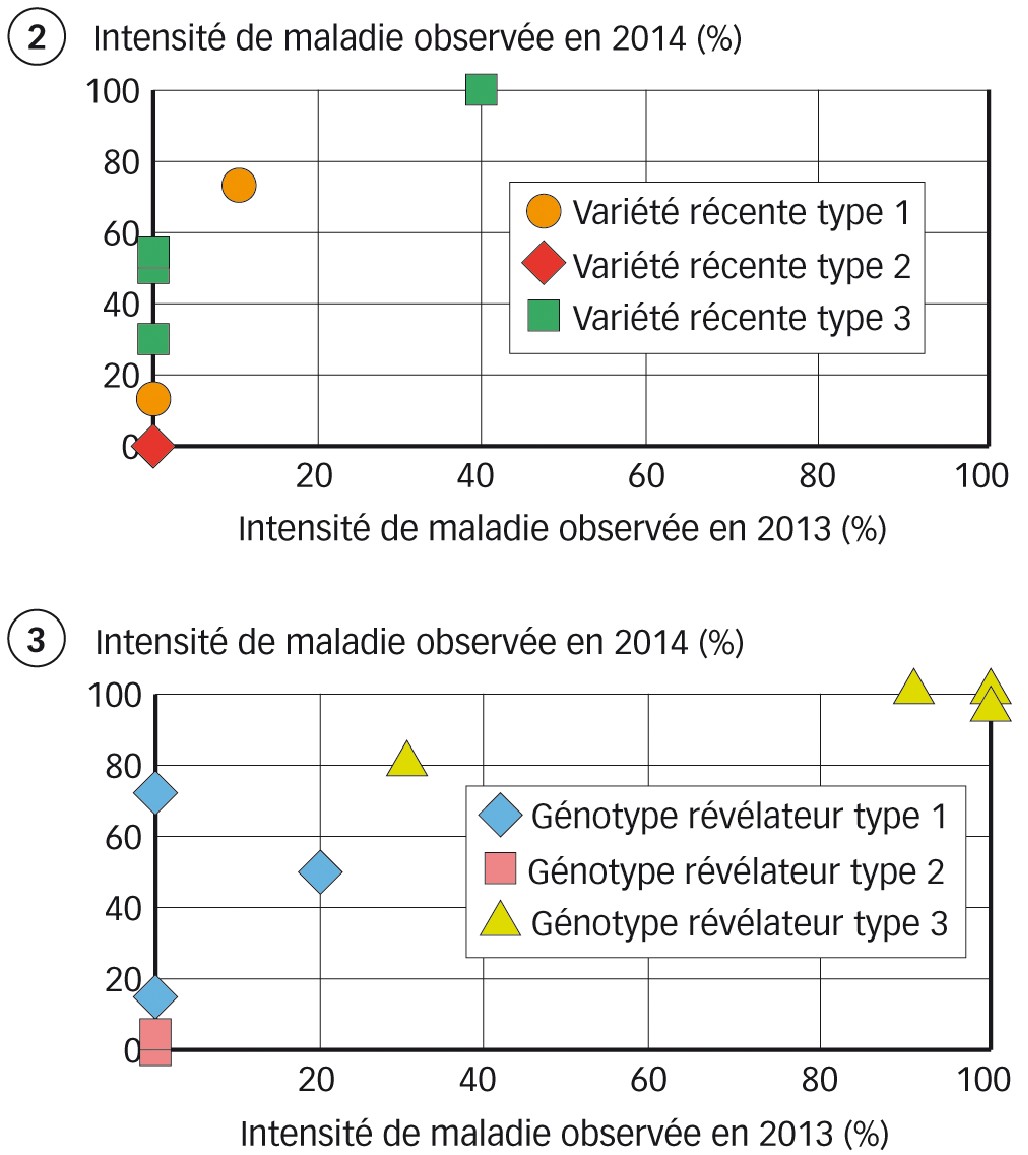
Figure 2 : exemples de variétés de blé récentes.
Figure 3 : exemples de génotypes révélateurs.
Un point = une variété sur un site expérimental de coop. Le comportement des variétés peut différer selon les lieux et évoluer selon les années (type 1, 2 ou 3). Les causes d'instabilité peuvent être liées à la différence de pression entre années, à une régionalisation des souches et/ou à leur évolution. Des variétés récentes conservent un comportement résistant sur tous les sites (variété de type 2).
Fig. 4 : Nombre de générations mensuelles théoriques pour les campagnes 2013, 2014 et 2015
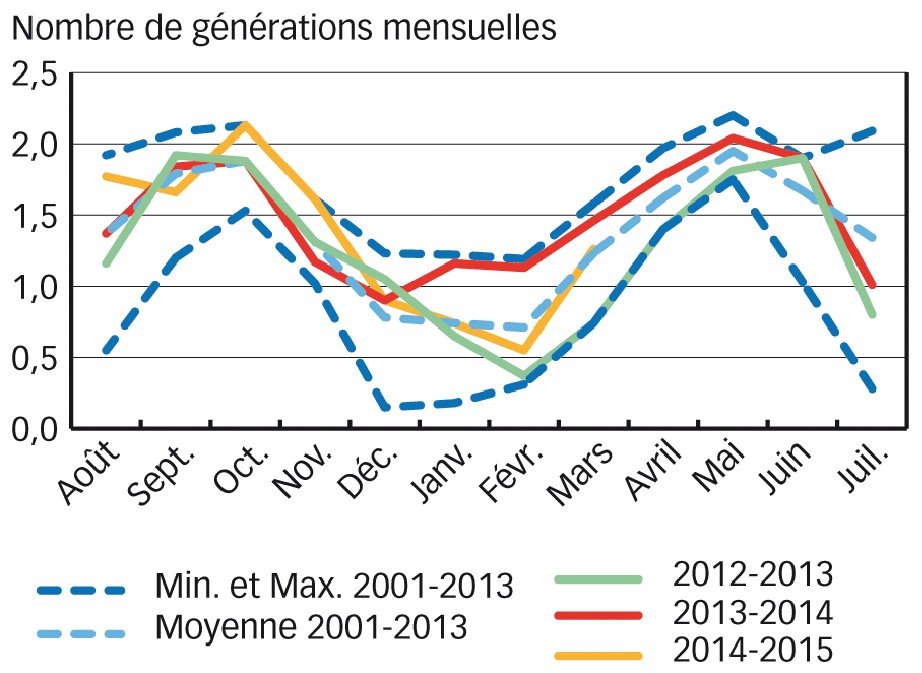
En 2013, les conditions printanières défavorables ont limité le développement de la rouille jaune en France. En revanche, les conditions hivernales et printanières de 2014, propices au développement du pathogène sur l'ensemble de la période, ont conduit à l'année épidémique la plus importante en surface et en intensité de ces quinze dernières années. En 2015, compte tenu de l'importance de l'inoculum et malgré un mois de février plutôt défavorable, la rouille jaune est à observer avec vigilance.
REMERCIEMENTS
Le projet FSOV (2012-2015) a financé l'étude des résistances variétales dans les essais menés par les coopératives et les obtenteurs partenaires ; l'étude sur l'évolution des races de rouille jaune a été financée en partie par le projet PURE (programme européen PCRD7) et par le contrat FSOV (2012-2015). Nous remercions toutes les personnes qui ont participé à la mise en place, au suivi et à l'analyse des essais. Nous remercions également Météo France qui a fourni l'ensemble des données météorologiques utilisées ici.

