 Imprimer
Imprimer
Mildiou sur feuille de pomme de terre. L'épidémie a été plus précoce et dommageable en 2014 qu'en 2013 partout en Europe. Inra Rennes-Le Rheu-R. Corbière
Depuis près de vingt ans, des chercheurs européens travaillant sur la pomme de terre échangent au sein du réseau EuroBlight. « Euro » comme européen, « Blight » comme potato blight, alias le mildiou, mais aussi early blight, alias l'alternaria.
Les activités d'EuroBlight
Des ateliers bisannuels permettent de déterminer les priorités de recherche et de vulgarisation et de les formuler dans des déclarations collectives.
Celles-ci peuvent servir de principes de base pour des actions conjointes et des collaborations internationales dans le but d'améliorer les stratégies de lutte intégrée (IPM, « integrated pest management »).
Des méthodes ont été harmonisées afin de fournir des outils pertinents aux chercheurs, généticiens, instituts techniques et firmes privées.
Le dernier atelier s'est tenu à Brasov, en Roumanie, en mai 2015. Il a réuni plus de cent participants de toutes les régions de l'Europe, Amérique du Sud, États-Unis, Israël et Chine.
Bilan épidémique
L'Europe du mildiou en 2013 et 2014
Les rencontres ont débuté par un résumé des épidémies des campagnes précédentes.
J. G. Hansen, du Danemark, et J. Leiminger, d'Allemagne, ont recueilli et présenté les données. L'épidémie de mildiou a été bien plus importante en 2014 qu'en 2013 dans toute l'Europe.
Elle a été nettement plus précoce en 2014, sauf dans les pays scandinaves.
Les écarts les plus nets entre les deux années ont été observés en France (début de contamination des parcelles, le 5 juillet en 2013, dès début mai en 2014), en Angleterre (1er août 2013, fin mai 2014) et en Écosse (1er septembre 2013, fin juin 2014).
Pour faire face à la maladie, de nombreux traitements ont été nécessaires en 2014. La présence d'oospores est relatée pour le Danemark, la Suède, la Pologne, la Lituanie et l'Estonie. Notons que les premiers échos de 2015 font état d'attaques relativement tardives et globalement modérées, notamment en France.
Du côté de l'alternaria
L'alternaria est signalé en 2013 dès le 15 juillet en Allemagne, mi-août à fin août aux Pays-Bas, pays scandinaves, pays baltes. En 2014, les signalements ont eu lieu le 5 juin en Allemagne, en juillet en France et aux Pays-Bas, fin juillet-début août en Belgique, Scandinavie et pays baltes.
Les épidémies sont souvent faibles même si plus sévères qu'ailleurs en Allemagne, Pays-Bas, Scandinavie et République tchèque. Des pertes de rendements de 7 à 15 % sont estimées pour l'Allemagne, les Pays-Bas, le Danemark et la Finlande.
Des gènes de résistance aux fongicides strobilurines ont été détectés. Il s'agit du gène F129 L sur A. solani aux Pays-Bas, en Allemagne et Suède, ainsi que du gène 143 A sur A. alternata en Belgique ainsi que, là encore, Pays-Bas, Allemagne et Suède.
Les populations de mildiou dans le monde
Évolution en Europe
D. Cooke (Écosse) a présenté le monitoring génotypique sur l'évolution des populations de Phytophthora infestans en Europe.
Pour déterminer le génotype de chaque isolat, une empreinte de chacun est faite sur une carte FTA ; ensuite une analyse des microsatellites de l'ADN (« multilocus SSRs ») est réalisée.
En 2014, il semble y avoir une augmentation des souches 6_A1 (Figure 1). En fait, ce résultat est biaisé : il y a eu, proportionnellement, davantage d'isolats issus de Grande-Bretagne en 2014 qu'en 2013. Or ce pays connaît une dominance des souches 6_A1 en 2014 comme en 2013.
La Figure 2 montre la répartition des souches en Europe en 2014. Les isolats 13_A2 et 6_A1 sont très fortement dominants dans les pays de l'ouest, sauf en Irlande où la lignée clonale 8_A1 reste bien présente.
Dans les pays de l'est et du nord, d'autres souches dominent. Dans ces régions, P. infestans survit essentiellement sous forme d'oospores provenant du croisement entre isolats de type A1 et du type A2, donnant de nouveaux génotypes issus de la reproduction sexuée ; l'inoculum primaire est généré localement et différent chaque année.
À l'inverse, en Europe de l'ouest, les populations se transmettent d'une année sur l'autre par reproduction asexuée, sous forme de clones. En France, une action d'épidémiosurveillance est en place(1).
Ailleurs dans le monde
Sur les autres continents, des réseaux de surveillance s'organisent.
E. Fry présente le réseau USA Blight, créé en 2009 à la suite d'une épidémie fulgurante très dommageable sur tomates de plein champ, mais aussi sur pommes de terre : cinq États avec de gros dégâts fin juin, 49 début août.
Un monitoring a été mis en place pour caractériser les populations de P. infestans responsables. Depuis 2012, un génotype particulier US-23 est devenu dominant aux États-Unis et a totalement remplacé le génotype US-8, majoritaire avant 2008(2).
Il semble que cette lignée clonale US-23 ait une préférence pour la tomate mais elle provoque aussi de graves dégâts sur pomme de terre. La recherche de résistance variétale est privilégiée.
I. Acuna (Chili) a présenté le réseau Tizon de dix pays d'Amérique latine créé en 2014. Diverses cultures sont sensibles : pomme de terre, tomate, narangille (tomates en arbre), coqueret du Pérou, pomme de terre créole. Un monitoring sur (= suivi de) la population de P. infestans est mis en place. La recherche de gènes de résistance est conduite sur de nouvelles espèces de Solanum sauvages. Greg Forbes a montré le travail de vulgarisation du CIP(3) dans plusieurs pays, notamment en Asie avec la création récente d'AsiaBlight.
Cycle biologique et épidémie
En France, souches et température
N. Mariette (Inra France) présente une étude, en lien avec le changement climatique, pour déterminer s'il existe une adaptation des souches à la température. Il compare des isolats d'origine géographiques et climatiques pour plusieurs traits d'agressivité (notamment taille des lésions, production et taille des spores) à quatre températures 10, 14,18, 24 °C(4).
Royaume-Uni : modèle et souches 6_A1
S. R. Dancey (UK) retravaille le modèle épidémiologique « Smith Period » et analyse, sur les années 2003-2013, les périodes où le modèle n'a pas anticipé l'arrivée de la maladie. L'étude prend en compte la fréquence des souches 6_A1 (environ 50 % de la population), 13_A2 (de l'ordre de 30 %), 8_A1 (10 %), les températures moyennes journalières, l'humidité relative et les variétés de pomme de terre.
Dans des tests sur folioles, en laboratoire à 10 °C avec 11 h Hr > 90 %, les infections sont plus importantes avec les isolats 6_A1 qu'avec les isolats 13_A2. Le phénomène s'amplifie à 14 °C, et il y a même des infections dès 6 h Hr > 90 % et 11 h à 80 % Hr avec les isolats 6_A1 testés.
Ces études montrent une agressivité plus forte de la lignée clonale 6_A1 que de la lignée clonale 13_A2 dans les conditions expérimentales. Une prise en compte de température et d'hygrométrie plus faibles dans les modèles est nécessaire.
Suède et Danemark : études sur les débuts d'épidémies
L'origine de l'inoculum primaire est étudiée : L. Sjöhlm (Suède) confirme le rôle dominant des oospores en Scandinavie mais attire aussi l'attention sur le rôle important des plants contaminés.
R. Naerstad (Danemark) rappelle l'importance des oospores dans le démarrage très précoce des épidémies qui sont alors difficiles à prévoir. Il présente l'étude d'un modèle tenant compte de l'humidité du sol et du piégeage de sporanges dans l'air.
Génotype et phénotype : résistance variétale
Pays-Bas : gènes et lutte raisonnée
G. Kessel (Pays-Bas) étudie le lien entre génotype, phénotype et stratégie de lutte raisonnée. Il cite les causes des difficultés pour combattre efficacement le mildiou : conditions climatiques très favorables à l'épidémie (voir en 2007 et 2014 en France), inoculum primaire important, évolution des populations de P. infestans.
La connaissance du génotype permet d'identifier les souches responsables. Elle devrait permettre, à l'avenir, de mieux comprendre la dynamique des populations due aux pressions de sélection exercées par les fongicides mais aussi par les variétés, etc.
Définir s'il existe des liens étroits entre le phénotype (caractéristiques biologiques) et le génotype (analyse de l'ADN) des isolats, serait très utile vu la rapidité de réalisation des analyses des ADN. Mais pour l'instant, les données sont limitées.
G. Kessel conclut son exposé sur le gros intérêt de la lutte intégrée avec introduction de gènes de résistance via cisgenèse.
Gènes empilés, cisgenèse et lutte intégrée aux Pays-Bas
Il est rejoint sur ce sujet par J. Vossen et T. Van den Bosch (Pays-Bas) qui mettent en valeur « la R'murie » , l'armurerie permise par les gènes R de résistance. Les auteurs insistent sur l'intérêt de l'empilement de gènes R de résistance spécifique via la cisgenèse associée à une lutte intégrée (IPM 2) avec un monitoring de virulences des souches pour détecter et suivre le contournement des gènes R.
Des résultats d'essais donnent une indication sur l'impact fongicide (Figure 3). Aux Pays-Bas, pour contrôler la maladie sur Désirée, à l'heure actuelle, il faut entre neuf et dix traitements à dose élevée, soit un IFT (indice de fréquence de traitement) de 12. Sur A 01, A15, A17, Bionica et Carolus, il suffit de neuf traitements à dose réduite donc un IFT de 3. Sur A10, A19, A16, A14 qui portent chacune deux à trois gènes R, la maladie est contrôlée sans aucun traitement.
En Écosse, le jeu des résistances
A. Lee (Écosse) cherche à comprendre comment la résistance variétale et la résistance aux fongicides sont liées, afin d'augmenter la durabilité du contrôle de P. infestans.
En effet le déploiement de variétés résistantes réduit la pression de sélection qui mène P. infestans, l'agent du mildiou, vers la résistance aux fongicides.
De même, le déploiement de fongicides réduit la pression de sélection qui mène P. infestans à contourner les gènes résistants des variétés de pomme de terre.
Production biologique
Variétés en Allemagne
K. Sieber (Allemagne) a évoqué un suivi de 150 variétés dans le but d'étudier des liens entre la résistance au mildiou et d'autres critères de qualité.
Il a mis un accent particulier sur l'intérêt de marqueurs génétiques pour repérer plus rapidement les gènes de résistances.
Éliciteurs en France
D. Andrivon (Inra France) a testé des éliciteurs pour lutter contre le mildiou à l'Inra de Ploudaniel. Selon ses travaux réalisés dans le cadre du projet européen CO-Free (« protection biologique » sans cuivre), certains éliciteurs présentent une efficacité partielle contre le mildiou. Mais toutes les variétés de pomme de terre ne réagissent pas de façon identique. La réponse de la plante déclenchée par les éliciteurs dépend de l'interaction entre éliciteur et plante.
Une bonne conduite de lutte (IPM) sur cultures « conventionnelles » ou biologique devrait prendre en compte toutes les formes de résistance de la plante : l'effet des gènes impliqués dans les mécanismes généraux de défense de la plante pourrait renforcer les gènes de résistance R ou ralentir leur contournement par des souches virulentes de P. infestans.
Produits antimildiou
Nouveautés fongicides
Des présentations ont porté sur plusieurs fongicides : Banjo Forte (dimétomorphe + fluaziname) par Adama, Consento (fénamidone + propamocarbe), Infinito (fluopicolide + propamocarbe) par Bayer. UPL a exposé l'intérêt des multisites comme le mancozèbe pour une lutte intégrée.
Des études en Russie sur Zeroxé à base de nanoparticules d'argent ont montré une efficacité en laboratoire sur de nombreux agents fongiques et bactériens de la pomme de terre.
Réglementation de ces produits
Une présentation très complète de l'évolution de la législation européenne a souligné le risque de difficulté de lutte en cas de retrait de certaines matières actives.
Le tableau avec l'efficacité et les caractéristiques des fongicides a été complété (il est à consulter sur le site internet d'EuroBlight(5)).
Mieux diagnostiquer l'alternaria et les Alternaria spp.
Suivi des deux espèces
H. Hausladen, B. Adolf et J. Leiminger, de l'université de Munich, ont fait plusieurs exposés de leurs travaux. Ils rappellent la présence des deux espèces en Europe Alternaria solani (avec deux génotypes : BI et BII) et Alternaria alternata.
Un des objectifs du groupe Alternaria est d'harmoniser les méthodes d'identification des espèces. Pour les prélèvements et la détermination microscopique, un « ring test » (test entre plusieurs laboratoires) a été mis en place ; les deux espèces sporulent en conditions différentes, une proposition de méthode d'identification est faite.
Symptômes « type alternaria » sans que les Alternaria spp. soient la cause
Des essais en serre et au champ conduits par B. Adolf et J. Leiminger en Allemagne sur Alternaria alternata sur la variété Markies ont montré l'apparition brutale de symptômes « type A. alternata », mais les analyses par qPCR ont démontré l'absence du pathogène sur 75 % des nécroses.
Les auteurs soulignent que (monitoring de 2005 à 2014) si A. alternata est presque toujours détecté, et A. solani dans 60 % des feuilles échantillonnées, ils ne sont pas toujours la cause des symptômes. Ces derniers pourraient être dus à des phénomènes physiologiques ou de phytotoxicité (ozone...). J. Spoelder constate d'ailleurs aussi aux Pays-Bas une grande proportion de symptômes « type alternaria » qui ne sont en fait pas dus à des Alternaria spp.
Alternaria et résistance aux strobilurines
Un gène de résistance par espèce
Selon des études conduites sur les résistances aux QoI, le gène F129L induit une résistance « partielle » (sensibilité réduite) d'isolats d'A. solani aux strobilurines, le gène G 143 A entraîne un fort niveau de résistance d'isolats d'A. alternata à ces matières actives.
Suivi sur A. solani
Le monitoring conduit de 2005 à 2014 en Allemagne, sur la mutation F 129L d'A. solani donne pour résultats :
- de 2005 à 2008, peu d'isolats et pas de traitement strobilurine, aucune mutation détectée ;
- de 2009 à 2012, progression d'usage des strobilurines, augmentation du nombre de sites avec mutants : 5,1 % (2009), 15,3 % (2010), 56,5 % (2011), 17 % (2012) ;
- en 2013 et 2014, sans traitement, la résistance progresse encore : 37,7 % et 32,2 %.
Il faut constater que la EC 50 (concentration qui assure 50 % de l'effet) des souches sans gène F129L reste stable. En revanche, celle des souches avec le gène de résistance augmente : trois à cinq fois la dose de matière active de référence en 2009 ; en 2012, quinze fois la dose, en 2014, 102 fois...
Suivi sur A. alternata
Pour le gène G 143 A d'A. alternata, le monitoring mis en place dès 2005 révèle les premières détections dès 2006, puis de 2009 à 2014, 30 % à 70 % des sites sont « contaminés ». L'étude des EC 50 montre un très haut niveau de résistance des souches à la matière active.
Lutte fongicide contre l'alternaria
Conseils généraux
Afin d'éviter des problèmes au champ, les chercheurs conseillent de limiter l'usage des fongicides à base de strobilurines, d'utiliser des fongicides efficaces à pleine dose, avec le premier traitement sept semaines après émergence et les suivants en fonction des conditions climatiques et de la progression des risques de la maladie.
Suivi en Suède
E. Liljeroth (Suède) aborde l'alternaria sous plusieurs aspects : différences de sensibilité variétale, nuisibilité conséquente nécessitant plusieurs traitements. Elle analyse la fréquence du gène F129L d'A. solani. Cette fréquence est importante dans le génotype GII mais plus faible dans le génotype GI. Des essais de produits seuls ou en programme sont conduits en quatre traitements avec Revus top (mandipropamide +difénoconazole), Signum (boscalid +pyraclostrobine) et Amistar (azoxystrobine).
Dans certains essais, l'azoxytrobine seule est du niveau du témoin, des isolements monospores montrent que cette utilisation entraîne une très nette augmentation de la population avec le gène F 129 L.
Cela peut expliquer que dans certaines parcelles de producteurs, la population d'Alternaria sp. est composée de près de 100 % d'isolats GII, portant le gène F 129 L.
Essais au Danemark
B. J. Nilsen (Danemark) a présenté des résultats d'essais fongicides en contamination artificielle (grains d'orges contaminés au sol). Sont testés Amistar, Signum, Revus +, seuls ou en programme, avec en moyenne quatre traitements. Les gains de rendement sont de l'ordre de 6,5 % et la date du premier traitement se situe à cinq à six semaines après émergence.
Suivi en Flandres belges
Un programme prévu sur cinq ans
P. Vanhaverbeke présente les premiers résultats du programme d'étude 2013/2018 de la station de recherche du PCA en Flandres belges. Un monitoring d'isolats est conduit hebdomadairement sur environ 150 champs.
En 2013, la maladie a été très faible du 1er juillet à fin août avec seulement A. alternata puis, de fin août à fin juillet, les deux espèces sont présentes.
En 2014, avec une épidémie plus sévère, seul A. alternata est détecté en juillet, puis les deux espèces sont présentes avec une dominante d'A. solani.
En 2013, il semble que la maladie soit due essentiellement à la sénescence. En présence d'A. solani, les nécroses sont plus précoces.
Premiers enseignements
P. Vanhaverbeke conclut que l'état physiologique et le stade de la culture sont au moins ou même plus importants que les conditions climatiques et la présence de l'inoculum.
La sénescence (ou la maturation des tubercules) semble être la condition de développement de la maladie dans une culture non stressée.
La dispersion de la maladie est plus lente que pour le mildiou, même si une feuille peut être détruite en huit à dix jours. De sévères attaques sur une culture sénescente n'entraînent pas de perte significative de rendement en Belgique. Toutefois, un modèle épidémiologique est en cours de construction, il prend en compte de nombreuses données climatiques mais aussi la physiologie de la plante.
Les modules pour les étapes du cycle : germination, croissance des taches, sporulation, dispersion sont bien avancés, alors que les modules sur l'infection et le potentiel de sporulation sont encore à améliorer. Côté fongicides, plusieurs fongicides ont été ajoutés au tableau consultable sur le site.
Conclusion
Du côté du mildiou
Pour Phytophthora infestans, les conditions climatiques sont l'élément essentiel au développement de l'épidémie. Il est toutefois indispensable de bien connaître l'évolution des populations du pathogène. Grâce au réseau EuroBlight, une base de données sur les isolats permet de comparer la structure et l'évolution des populations de P. infestans en Europe.
Par ailleurs, établir des liens entre génotype et phénotype des isolats serait très utile. Les « nouvelles souches » présenteraient des différences biologiques avec les « anciennes ». La résistance variétale est un élément très fortement pris en compte dans la lutte.
Les Alternaria spp.
Concernant les Alternaria spp., l'harmonisation des méthodes de détection et de détermination des espèces pathogènes, des expérimentations en cours sont disponibles sur le site d'EuroBlight.
Dans l'ouest de l'Europe, l'importance de la maladie est probablement surévaluée, beaucoup de symptômes ne sont pas dus à ces pathogènes. L'impact sur le rendement est plus élevé « à l'est » comme en Allemagne.
L'état physiologique de la plante est aussi important, voire plus, que les conditions climatiques, d'où probablement l'absence de modèle efficace pour l'instant ; mais plusieurs chercheurs y travaillent.
En matière de lutte, les traitements doivent être faits tôt, dès les premiers symptômes, voire avant. La résistance aux QoI est bien présente dans plusieurs pays même si des situations de non-efficacité au champ ne sont pas relevées.
(1), (2) et (4) Voir « Mildiou en France : P. infestans sous surveillance », p. 30). (3) Centre international de la pomme de terre. ((5) Site EuroBlight : http://euroblight .net.
Fig. 1 : Mildiou en Europe, en 2013 puis 2014

Source : D. Cooke - EuroBlight
Identité des génotypes sur 712 échantillons (26 pays) en 2013, et 1 552 échantillons de 24 pays en 2014.
Fig. 2 : Mildiou en Europe : où sont les génotypes ?
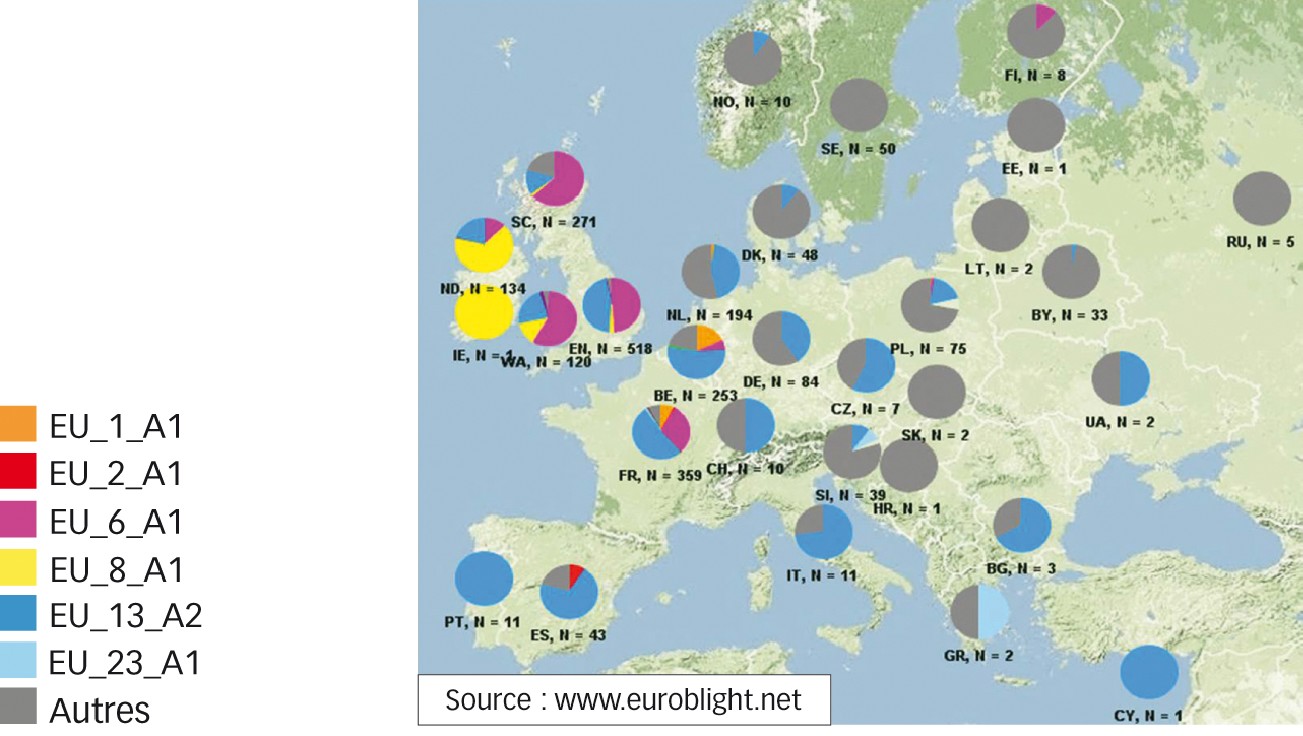
Répartition géographique des principaux génotypes européens de P. infestans collectés en 2013 et 2014.
Fig. 3 : Intérêt de l'empilement de gènes de résistance pour diminuer l'utilisation des fongicides dans le cadre de l'IPM2.0

Source : J. Vossen & T. Van den Bosch
Entre la variété Désirée, dont la résistance partielle s'est érodée, et les variétés ayant plusieurs gènes de résistance efficaces, il existe des variétés à traiter à faible dose. IPM = protection intégrée.

