 Imprimer
Imprimer
Photo : M. H. Balesdent
 par test cotylédonaire à l'aide de souches caractérisées (Pinochet et al., 2009). À droite, données de superficies cumulées pour chacun des gènes (variétés avec Rlm7 seul ou associé avec Rlm3, variétés avec Rlm3 seul ou associé à Rlm7).")
Fig. 1 : Évolution des surfaces de colza cultivées en France avec les variétés portant les gènes de résistance Rlm7 et Rlm3 Les surfaces cultivées avec les différentes variétés de colza sont issues du Panel Agriscan, qui repose sur une enquête trimestrielle des pratiques en grandes cultures de 9 500 agriculteurs. Les variétés ont été caractérisées pour la présence de Rlm3, Rlm7 ou les deux (à gauche) par test cotylédonaire à l'aide de souches caractérisées (Pinochet et al., 2009). À droite, données de superficies cumulées pour chacun des gènes (variétés avec Rlm7 seul ou associé avec Rlm3, variétés avec Rlm3 seul ou associé à Rlm7).
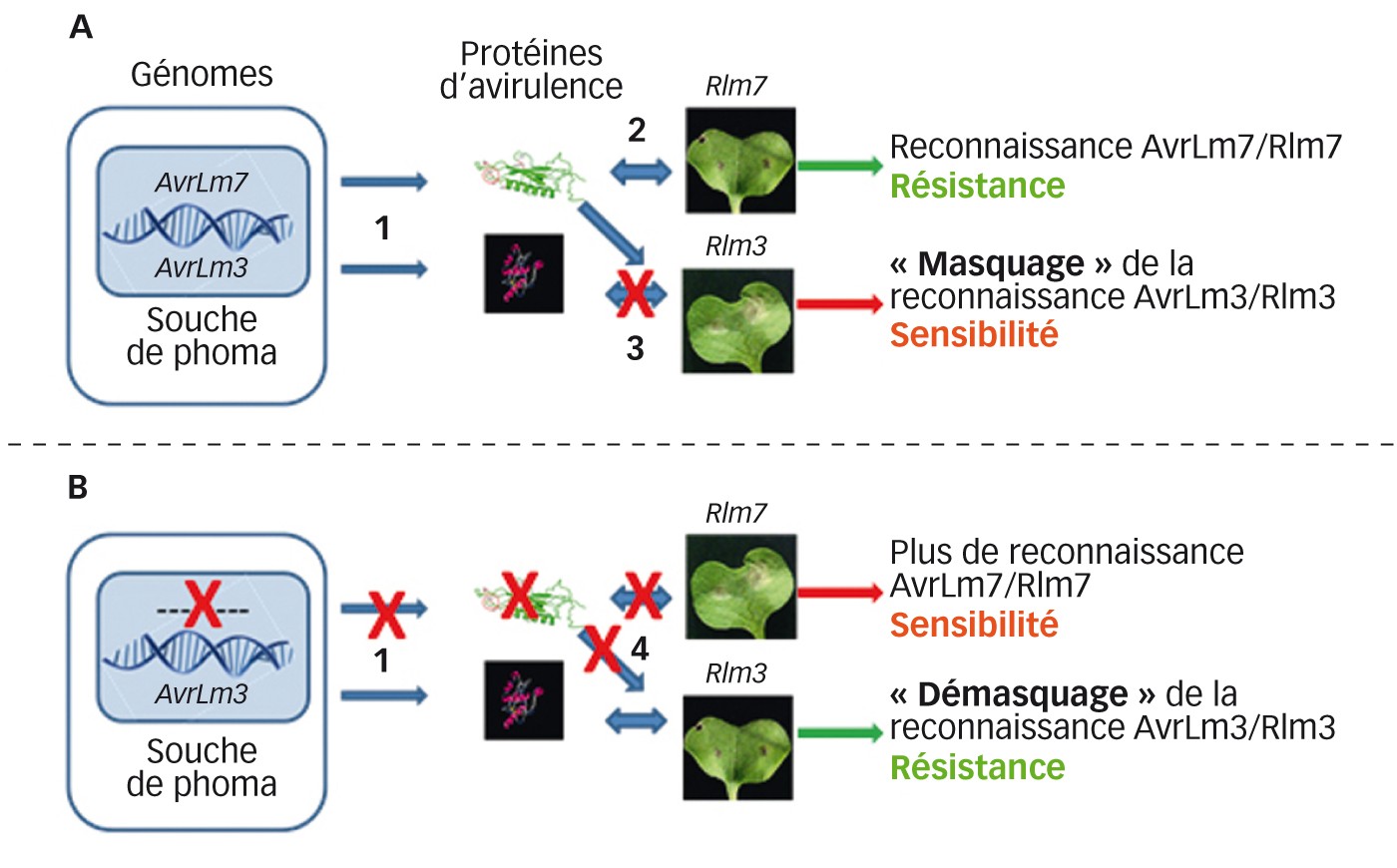

Fig. 3 : Évolution des fréquences de souches virulentes sur Rlm7 en France depuis 2009 et état des lieux des fréquences de souches « double virulentes » sur Rlm3 et Rlm7 en 2017 Chaque diagramme circulaire représente un site échantillonné. Les proportions de souches virulentes sur Rlm7 sont représentées en rouge. Sur la carte « 2017b », les proportions de souches « double virulentes » sur Rlm3 et Rlm7 sont représentées en orange, comparées aux souches représentées en bleu, qui correspondent à la proportion de souches qui peuvent être contrôlées soit par Rlm7, soit par Rlm3. Les cercles entourés de noir correspondent à des parcelles expérimentales avec apport d'inoculum local renforcé.
. La protéine AvrLm3 est produite (1) mais n'est plus reconnue par Rlm3. Les variétés Rlm3 sont donc également sensibles à ce type de souche. B. Souches du groupe 2 : le gène AvrLm7 porte une mutation ponctuelle. La protéine AvrLm7 mutée () est produite et sécrétée (1), mais n'est plus reconnue par la protéine de résistance Rlm7 (2), et la variété Rlm7 est sensible à cette souche. Mais cette mutation ne modifie pas la capacité d'AvrLm7 à empêcher la reconnaissance entre AvrLm3 et Rlm3 (3). Malgré la production de la protéine d'avirulence AvrLm3, les génotypes de colza portant Rlm3 sont donc sensibles à ce type de souche.")
Fig. 4 : Événements moléculaires chez les souches « double virulentes » vis-à-vis des gènes de résistance Rlm3 et Rlm7 A. Souches du groupe 1 : le gène AvrLm7 est inactivé ou délété, la souche ne produit plus la protéine d'avirulence correspondante. De ce fait, les variétés Rlm7 sont sensibles à cette souche. Le gène AvrLm3 porte une mutation ponctuelle (). La protéine AvrLm3 est produite (1) mais n'est plus reconnue par Rlm3. Les variétés Rlm3 sont donc également sensibles à ce type de souche. B. Souches du groupe 2 : le gène AvrLm7 porte une mutation ponctuelle. La protéine AvrLm7 mutée () est produite et sécrétée (1), mais n'est plus reconnue par la protéine de résistance Rlm7 (2), et la variété Rlm7 est sensible à cette souche. Mais cette mutation ne modifie pas la capacité d'AvrLm7 à empêcher la reconnaissance entre AvrLm3 et Rlm3 (3). Malgré la production de la protéine d'avirulence AvrLm3, les génotypes de colza portant Rlm3 sont donc sensibles à ce type de souche.
 ou groupe 2 (mutation ponctuelle dans AvrLm7 et allèle avirulent chez AvrLm3).")
Fig. 5 : Caractérisation moléculaire des souches de phoma « double virulentes » présentes en France Par analyse moléculaire des gènes AvrLm3 et AvrLm7, les souches sont classées en « groupe 1 » (AvrLm7 inactivé, allèle virulent pour AvrLm3) ou groupe 2 (mutation ponctuelle dans AvrLm7 et allèle avirulent chez AvrLm3).
. Les courbes représentent les ajustements aux données (courbes polynomiales d'ordre 3, r2 = 0,931 et 0,998, respectivement, pour Rlm1 et Rlm7).")
Fig. 6 : Évolution des souches virulentes sur Rlm1 et sur Rlm7 après introduction des gènes de résistance dans les variétés de colza en France Les années initiales ont été choisies pour correspondre à une même fréquence initiale de souches virulentes (1995 pour Rlm1, 2010 pour Rlm7). Les courbes représentent les ajustements aux données (courbes polynomiales d'ordre 3, r2 = 0,931 et 0,998, respectivement, pour Rlm1 et Rlm7).
La nécrose du collet, ou « phoma », provoquée par le champignon Leptosphaeria maculans, est l'une des principales maladies cryptogamiques du colza. Ces dernières années, la maladie est restée discrète, grâce à l'amélioration génétique des variétés de colza pour leur résistance au phoma.
Résistances génétiques du colza
Partielle ou totale
Deux types de résistance au phoma sont décrits chez le colza (Delourme et al., 2006) : d'une part les résistances quantitatives, efficaces vis-à-vis de toutes les souches de l'agent pathogène, mais n'apportant qu'un niveau partiel de résistance (les plantes peuvent être infectées, mais l'intensité de la nécrose au collet reste réduite et a un faible impact sur le rendement) ; d'autre part, les résistances contrôlées par des gènes majeurs de résistance (gènes Rlm), qui bloquent l'infection primaire, et donc apportent une protection totale vis-à-vis de la fraction de la population pathogène portant les allèles avirulents des gènes d'avirulence (AvrLm) correspondants (Balesdent et al., 2015 ; Rouxel et Balesdent, 2017).
Gènes de résistance Rlm3 et Rlm7
Les hybrides de colza portant le gène de résistance spécifique Rlm7 ont connu un succès croissant depuis 15 ans. En effet, face à une population de phoma composée à plus de 99,5 % de souches avirulentes sur Rlm7 en 2000 (Balesdent et al., 2015), ces hybrides portaient une résistance particulièrement efficace. Ainsi, le pourcentage estimé de surfaces de colza cultivées avec des variétés portant Rlm7 est passé de 25 à 60 % entre 2011 et 2013 (Figure 1).
En 2013, une caractérisation approfondie des populations de phoma avait montré une évolution de celles-ci depuis 2000 : des souches capables de contourner la résistance Rlm7 étaient détectables dans onze des douze sites échantillonnés, avec une fréquence moyenne de 19,5 % sur le territoire français (Balesdent et al., 2015). Lors de cette analyse, ou dans le cadre de travaux plus larges sur des populations mondiales, nous avions démontré une relation inverse quasi systématique entre l'aptitude des souches à attaquer les variétés Rlm7 et celle à attaquer les variétés Rlm3 : les souches contournant Rlm7 étaient en très grande majorité avirulentes sur les variétés portant le gène Rlm3, et inversement (Plissonneau et al., 2015, 2016). Par un mécanisme moléculaire non encore élucidé, la présence du gène AvrLm7 dans une souche suffit pour masquer la reconnaissance entre Rlm3 et AvrLm3, mais si le gène AvrLm7 est inactivé, ce masquage disparaît et la reconnaissance AvrLm3-Rlm3 peut avoir lieu, induisant la résistance due à Rlm3 (Figure 2 p. 32). Ce mécanisme offrait des perspectives intéressantes de redéploiement du gène de résistance Rlm3 (Plissonneau et al., 2015), alors qu'il avait été considéré comme totalement inefficace en Europe dans les années 2000 (Huang et al., 2010).
Toutefois, dès 2013, quelques souches capables de contourner à la fois les résistances Rlm3 et Rlm7 (souches dites « double virulentes » ou « a3a7 ») étaient détectées dans près de la moitié des sites échantillonnés, bien qu'à des fréquences très faibles (moins de 0,5 % des souches à l'échelle nationale).
Surveillance des populations de phoma depuis 2013
Isolement de souches de 2013 à 2017
Pour faire suite aux analyses réalisées à l'échelle nationale en 2000-2001, puis 2009-2010, nous avons réalisé un suivi régulier des populations de phoma de 2013 à 2017. Pour cela, des variétés « pièges », ne possédant ni Rlm3 ni Rlm7, ont été semées en parcelles d'agriculteurs dans des sites correspondant aux principales régions de culture du colza. Quelques sites expérimentaux, dans lesquels des résidus de colza infectés récoltés localement ont été apportés pour renforcer l'inoculum (Brun et al., 2009), ont également été caractérisés. Pour chaque site, des populations de phoma (50 à 100 souches par sites) ont été isolées à l'automne à partir de « macules » formées sur les feuilles des variétés pièges. Ces souches ont ensuite été caractérisées pour leur virulence après inoculation en conditions contrôlées sur des variétés possédant Rlm3, Rlm7 ou aucun de ces gènes (Balesdent et al., 2015 ; Leflon et al., 2015). Au total, 4 073 souches ont été isolées et caractérisées, provenant de neuf à dix-sept sites selon les années (tableau p. 32).
Évolution des fréquences de souches virulentes sur Rlm7
La progression des souches virulentes sur Rlm7 est régulière et atteint en moyenne 50 % des souches en 2017 (tableau ; somme des phénotypes A3a7 et a3a7). Le contournement de Rlm7 est donc effectif et généralisé (Figure 3 p. 33). Il existe toutefois des différences selon les sites, y compris dans les sites d'expérimentation avec inoculum renforcé. Ainsi, les sites situés en région de faible production de colza sont ceux pour lesquels la proportion de souches virulentes reste faible, comme la Bretagne de 2013 (0 %) à 2015 (1,5 %) ; l'Isère en 2014 (6,7 %), ou la Haute-Garonne en 2017 avec moins de 10 % de souches virulentes.
Des sites assez proches peuvent présenter des fréquences de souches virulentes significativement différentes pour une même campagne. Par exemple, en 2017, les populations isolées de Précy-sur-Vrin (Yonne) et Puits (Côte-d'Or), distants de seulement 86 km, diffèrent significativement (test non paramétrique de Dunn, p = 0,006) pour leurs pourcentages de souches virulentes sur Rlm7 (34,7 % et 82,7 %, respectivement). Ce résultat suggère donc des variations locales, qui peuvent être liées aux variétés dominantes cultivées localement ou à des différences locales de pratiques culturales ou de situation agronomique.
Un contournement simultané de Rlm3 et Rlm7 amorcé en 2017
Entre 2009-2010 et 2017, les populations de phoma restent en grande majorité soit avirulentes sur Rlm7 (phénotype « a3A7 », tableau), soit avirulentes sur Rlm3 (phénotype « A3a7 »), puisque ces deux phénotypes cumulés représentent respectivement 99,6 %, 99,0 %, 98,8 % et 95,0 % des souches en 2013, 2014, 2015 et 2017. Des souches capables d'attaquer les variétés cumulant les deux gènes (souches « a3a7 ») sont rares et détectées dans moins de la moitié des sites en 2013, 2014 et 2015, mais sont détectées dans 14 sites sur 17 en 2017. Plus inquiétant, en 2017, de telles souches sont détectées à plus de 5 % dans cinq sites, pour atteindre autour de 20 % dans deux sites proches (Figure 3). L'accroissement de la proportion de souches a3a7 et donc le contournement simultané de Rlm3 et Rlm7, est clairement amorcé à l'automne 2017. Avec en moyenne 5 % de souches a3a7 en 2017, nous nous retrouvons, vis-à-vis des variétés pyramidant Rlm3 et Rlm7, au même niveau de contournement qu'en 2009-2010 vis-à-vis de Rlm7 seul (Figure 3).
Mécanismes moléculaires de contournement
Deux groupes de souches double virulentes
Les gènes d'avirulence AvrLm3 et AvrLm7 ont été clonés, ce qui permet d'étudier par quels mécanismes moléculaires les souches contournent simultanément les deux résistances. Dans une étude portant sur une collection mondiale de plus de 500 souches, nous avons montré que les souches a3a7 appartiennent à deux grands groupes (Plissonneau et al., 2017 ; Figure 4 p. 34).
Groupe 1 : perte et mutation de gènes
Les souches du groupe 1 ont perdu le gène AvrLm7 (ou en possèdent une version très altérée), ce qui leur confère la virulence sur Rlm7. Elles possèdent par ailleurs une version mutée du gène AvrLm3, conférant un phénotype virulent sur Rlm3. À ce jour, quatre allèles virulents différents d'AvrLm3, avec des mutations ponctuelles différentes, ont été identifiés dans ce groupe de souches. Tous ces allèles permettent néanmoins la synthèse d'une protéine « fonctionnelle », c'est-à-dire toujours secrétée hors de la cellule du champignon et n'ayant a priori pas de modification de structure tridimensionnelle. Ces versions mutées de la protéine AvrLm3 ont perdu leur fonction de reconnaissance par Rlm3 mais ont probablement conservé leur fonction « effectrice(1) » qui caractérise les gènes d'avirulence (Figure 4A) (Plissonneau et al., 2017).
Groupe 2 : mutations ponctuelles d'un seul gène
Les souches a3a7 du groupe 2 sont caractérisées par de simples mutations ponctuelles dans AvrLm7 (Figure 4B), ne modifiant qu'un petit nombre d'acides aminés de la protéine d'avirulence. Là encore, ces mutations n'ont pas d'impact majeur sur la structure de la protéine AvrLm7. Chez ces souches, le gène AvrLm3 n'est pas différent des versions présentes chez les souches de phénotype A3a7, avirulentes sur Rlm3. Nous en concluons que ces simples mutations ponctuelles dans AvrLm7 suffisent à empêcher la reconnaissance AvrLm7-Rlm7, et donc à rendre la souche virulente sur Rlm7, sans altérer le masquage de la reconnaissance AvrLm3-Rlm3 par la protéine AvrLm7, ce qui maintient le phénotype virulent de ces souches sur Rlm3. De telles mutations ponctuelles dans AvrLm7 sont particulièrement parcimonieuses pour le champignon puisqu'elles permettent, via une seule mutation, de contourner simultanément deux gènes de résistance, tout en maintenant la production des deux protéines effectrices AvrLm3 et AvrLm7 et donc de maintenir un bon niveau d'agressivité chez ces souches (Plissonneau et al., 2017).
Souches double virulentes présentes en France
Pour identifier le type de souches double virulentes présentes en France, les gènes AvrLm7 et AvrLm3 ont été amplifiés par PCR et les produits PCR séquencés chez 29 souches a3a7 isolées en France entre 2007 et 2014 : dix souches issues des échantillonnages sur les variétés pièges, et dix-neuf souches isolées spécifiquement sur des variétés avec Rlm7, pour accroître l'effectif analysé. En 2007 et 2008, peu de souches a3a7 sont disponibles et toutes appartiennent au groupe 1 (Figure 5 p. 34). À partir de 2012, des souches des groupes 1 et 2 sont détectées et celles du groupe 2 sont majoritaires chaque année. Une très grande diversité d'événements de mutations est détectée. Les souches du groupe 2 présentent huit événements différents, combinant les huit isoformes distinctes d'AvrLm7 et deux isoformes d'AvrLm3. Chez les souches du groupe 1, où AvrLm7 est absent, quatre isoformes différentes ont été identifiées pour AvrLm3. Pour un même lieu et une même année de prélèvement, les souches a3a7 étudiées peuvent toutes présenter des mutations différentes. Par exemple, dans l'Indre en 2014, les cinq souches a3a7 étudiées montrent cinq combinaisons de mutations différentes. Ce résultat est tout à fait comparable à ce que nous avions observé expérimentalement en 2006-2008 à Grignon, où sur une même micro-parcelle soumise à forte pression de sélection Rlm7, les souches contournant Rlm7 présentaient une extrême diversité d'événements de mutations (Daverdin et al., 2012). Inversement, un même événement peut être retrouvé dans plusieurs sites ou plusieurs années d'échantillonnage.
Vers une stratégie d'utilisation des résistances variétales
Un contournement progressif de Rlm7
L'utilisation massive de la résistance Rlm7 en France depuis 15 ans a conduit progressivement à un accroissement de la proportion de souches contournant cette résistance. Aujourd'hui, Rlm7 est clairement contourné dans quasiment toutes les régions échantillonnées. Ce contournement a été cependant beaucoup plus lent que ce qui avait été observé historiquement lors de l'utilisation de Rlm1 dans les années 1990 où, en seulement trois saisons culturales, des fréquences de souches virulentes sur Rlm1 bien supérieures à celles observées aujourd'hui pour Rlm7 avaient été obtenues (Rouxel et al., 2003 ; Figure 6 p. 34).
Cette relative durabilité est probablement attribuable au niveau de résistance quantitative satisfaisant des cultivars portant Rlm7 et au mécanisme d'interaction négative entre AvrLm3 et AvrLm7. En effet, la proportion de variétés cultivées avec le gène Rlm3 a été non négligeable pendant de nombreuses années en France. Ainsi, 35 % des variétés inscrites entre 2001 et 2007 portaient le gène de résistance Rlm3 (Pinochet et al., 2009). En 2011, plus d'un tiers de la sole colza était cultivé avec des variétés portant Rlm3 (Figure 1). Ces génotypes ont pu ralentir la progression des souches virulentes sur Rlm7 en contre-sélectionnant chaque année les souches ayant surmonté la résistance Rlm7, de phénotype A3a7.
Un pyramidage favorable à la sélection de souches double virulentes
Or le profil de déploiement des résistances variétales en France a largement évolué depuis 2011, avec trois faits marquants :
- l'utilisation massive et croissante du gène Rlm7, atteignant près de 60 % de la sole colza à partir de 2013 ;
- en parallèle, une baisse des surfaces portant le gène Rlm3 « seul » (= non associé à Rlm7), avec dès 2014 moins de 10 % des surfaces cultivées avec Rlm3 seul ;
- l'apparition dans le paysage variétal de variétés pyramidant Rlm3 et Rlm7, culminant à 28 % des surfaces à l'automne 2013 (Figure 1).
Toutes ces conditions ont été particulièrement favorables à la sélection de souches a3a7, les seules capables d'attaquer les variétés possédant à la fois Rlm3 et Rlm7, selon le scénario suivant : l'utilisation massive de Rlm7 associée à la diminution de la contre-sélection due à Rlm3 a accéléré l'apparition des souches virulentes sur Rlm7 ; parmi celles-ci, celles de phénotype a3a7 ont à leur tour été sélectionnées sur les variétés pyramidant Rlm3 et Rlm7, tout en sélectionnant parmi celles-ci les moins affectées par les événements de contournement, c'est-à-dire les souches a3a7 du groupe 2. Ainsi, le pyramidage de Rlm3 et Rlm7 a pu contribuer à sélectionner les événements de mutation les plus favorables au champignon, puisque les souches a3a7 du groupe 2 sont capables de contourner les deux résistances grâce à une unique mutation ponctuelle n'altérant pas la structure des protéines AvrLm3 et AvrLm7 produites (Plissonneau et al., 2017 ; Figure 4). Certaines régions présentent déjà une proportion importante de souches a3a7, qui ne sont donc plus contrôlées ni par Rlm3 ni par Rlm7.
Afin de ne pas détruire définitivement le potentiel de ces deux gènes de résistance, à court terme dans ces régions et à plus long terme sur l'ensemble du territoire, il devient urgent de maintenir dans les rotations des proportions élevées de variétés sans Rlm7 mais à niveau élevé de résistance quantitative, de favoriser l'alternance des variétés Rlm3 et Rlm7 plutôt que de les pyramider et enfin de diversifier, dans la mesure du possible, le déploiement de nouveaux gènes de résistance spécifiques efficaces.
(1) Pour effectuer leurs cycles infectieux sur leurs hôtes, les agents phytopathogènes secrètent un ensemble de molécules (petites protéines, toxines...) capables de manipuler la cellule végétale à leur avantage ou de contrecarrer les voies de défense des plantes. Indépendamment de leur fonction de reconnaissance par la plante-hôte, les protéines d'avirulence sont aussi des protéines effectrices contribuant au pouvoir pathogène.REMERCIEMENTS Ce travail a bénéficié d'un soutien financier de l'AFB (ex-Onema) (Projet « Icoscop », 2012-2015), de l'Inra (projets du métaprogramme SMaCH « K-masstec » et « MogerR »), du Labex Basc (Projet « Dynamic-Rlm7 »), et de Sofiproteol (Projet Faso « Ephicas » 2017-2020). Les auteurs remercient pour leur implication et leurs contributions aux échantillonnages de populations de phoma : Loïc Le Meur ; les coopératives du réseau InVivo (Axéréal, Cavac, Bonneval Beauce et Perche, EMC2, CapSeine, Noriap, La Dauphinoise et Agrial) ; l'équipe du laboratoire génétique et protection des cultures de Terres Inovia ; Magali Ermel, Patrick Vallée et Claude Domin (Igepp Le Rheu) et l'UE La Motte (Inra Le Rheu) ; ainsi que Pierre George (Innolea) et les partenaires du réseau d'expérimentation d'Ephicas : Euralis Semences, Limagrain Europe et Ragt.
RÉSUMÉ
CONTEXTE - Le gène de résistance Rlm7 a été très largement utilisé dans les variétés de colza depuis 15 ans. L'apparition de souches de phoma capables de contourner Rlm7 a été lente mais régulière et s'est accompagnée de la réapparition de souches avirulentes vis-à-vis du gène Rlm3, permettant ainsi de réutiliser ce gène comme source de résistance efficace. Récemment, des souches de phoma capables de contourner simultanément Rlm3 et Rlm7 ont été détectées sur plusieurs sites et parfois de façon non négligeable.
SURVEILLANCE - Quatre années de suivi intensif des populations de phoma en parcelles agricoles ou expérimentales en France permettent de dresser un état des lieux, en France, des populations de phoma pour les virulences 3 et 7. La relation négative entre phénotypes virulents sur Rlm3 et Rlm7 est quasi systématique à l'échelle du territoire, mais commence à s'éroder.
Des souches double virulentes apparaissent. Les résultats suggèrent qu'il est risqué de poursuivre le déploiement de variétés pyramidant Rlm3 et Rlm7.
MOTS-CLÉS - Colza, phoma, Leptosphaeria maculans, gènes de résistance Rlm3 et Rlm7.
POUR EN SAVOIR PLUS
CONTACT : marie-helene.balesdent@inra.fr
BIBLIOGRAPHIE : la bibliographie de cet article (six références) est disponible auprès de ses auteurs (contact ci-dessus).

