
 Imprimer
Imprimer
ph. M. Doumergue
En arboriculture, la sélection variétale est un très long travail. Pour construire aujourd'hui les variétés de demain, comment peut-on définir les « idéotypes variétaux » c'est-à-dire comment prioriser les principaux caractères-cibles à combiner ? Ces caractères, qui peuvent varier selon les conditions socioéconomiques (coût et disponibilité de main-d'œuvre), climatiques, etc., sont puisés dans une variabilité génétique que nous allons décrire. Mais il faut aussi tenir compte des interactions variétésenvironnement-conduite du verger. Comment ? À partir de l'exemple des interactions entre le développement du pommier et sa résistance aux bio-agresseurs, nous ouvrons quelques pistes pour concevoir des idéotypes multi-critères « bas intrants » en rassemblant des compétences multidisciplinaires et en utilisant la modélisation.
La performance agronomique d'un verger repose sur des choix qui se déclinent à deux échelles de temps (Lauri et al., 2009).
Les choix initiaux sont posés lors de l'implantation : variétés scions et porte-greffe, distances de plantation, orientation des rangs, système de palissage.
Viennent ensuite, chaque année, les choix culturaux : interventions de conduite (taille, arcure...), éclaircissage, apports d'engrais, d'eau et de traitements phytosanitaires. Ces pratiques génèrent des coûts financiers et environnementaux ainsi que, pour les produits phytosanitaires et d'éclaircissage, des risques potentiels pour la santé des consommateurs et arboriculteurs. Pour un verger de pommier adulte, le coût de main-d'œuvre lié aux interventions (récolte, taille, éclaircissage, extinction) représente 50 à 60 % des coûts totaux en moyenne (GRCETA de Basse-Durance, 2009, com. personnelle ; Deschênes, 2002). La protection phytosanitaire requiert 30 à 38 traitements selon les bassins de production (Sauphanor et al., 2009) face aux principaux bio-agresseurs : carpocapse, puceron cendré, tavelure, oïdium, feu bactérien, maladies de conservation, etc.
De plus, le changement climatique en cours accentue le risque parasitaire et risque de pénaliser des variétés jusqu'ici performantes.
Face à ces pressions croissantes sur les conditions d'une production fruitière performante, il faut rénover les pratiques en associant les compétences de généticiens et sélectionneurs, d'agronomes, pathologistes, physiologistes, modélisateurs, socio-économistes et représentants de la filière arboricole.
En particulier, la conception des idéotypes variétaux (notion définie dans l'encadré 1) doit être repensée par une approche globale replaçant la variété dans son environnement, soumise à des pratiques culturales (traitements et manipulations), au sein d'une interaction complexe à optimiser pour répondre aux enjeux économiques et environnementaux déjà cités.
Cet article tente de dégager quelques idées forces sous-tendant les recherches conduites à l'INRA et permettant d'aborder le triple niveau de complexité devant être pris en compte dans la construction d'idéotypes :
– combiner des allèles favorables de plusieurs gènes pour chaque caractère,
– combiner plusieurs caractères dont le déterminisme génétique ou la physiologie peuvent s'avérer antagonistes,
– rechercher les synergies favorables entre potentialités génétiques, effets environnementaux et pratiques culturales.
Nous nous baserons sur le pommier qui peut servir de modèle pour d'autres espèces fruitières.
Variabilité du pommier
Le pommier, originaire d'Asie centrale, serait apparu il y a des dizaines de millions d'années (Juniper, 2007). Plusieurs études ont montré la diversité génétique des espèces sauvages, surtout Malus sieversii (Leded.) en Asie centrale (Wiedow et al., 2004) et Malus sylvestris (L.) en Europe, et la possible hybridation avec le pommier domestique (Wagner et al., 2004). Un effort de collecte et de rassemblement des ressources génétiques de l'espèce a été fait à la station de Geneva (état de New York, USA) qui a récemment introduit de nouvelles accessions (Volk et al., 2005).
L'UMR GenHort à l'INRA d'Angers gère les ressources génétiques pour les fruits à pépins au niveau national. Sa base de données contient plus de 2 000 variétés provenant de nombreux vergers conservatoires répartis sur tout le territoire. Le site d'Angers abrite près de 1 400 variétés dont 800 font partie de la collection nationale en cours de constitution. Des descriptions sont réalisées sur les principaux caractères morphologiques et agronomiques (phénologie, qualité du fruit, résistance aux parasites). De plus, 137 accessions représentent des taxons botaniques de plusieurs espèces de Malus provenant de différents pays.
Variabilité de la qualité des fruits
La plupart des programmes passés d'amélioration génétique étaient ciblés sur les attentes de l'arboriculture « intensive » et la demande de la grande distribution. Ils tendaient à standardiser les produits de leur sélection : gros fruits très colorés, sans défaut cosmétique et à bonne valeur gustative. De plus, les fruits devaient être fermes, juteux et croquants et se conserver le plus longtemps possible.
Mais les collections abritent une très grande variabilité pour tous les attributs de qualité : calibre, coloration, teneurs en sucre et en acide, parfum, arôme, texture, etc.
Dans le cadre du projet européen HiDRAS (2003-2007; http://users.unimi.it/hidras/), les principaux caractères de qualité du fruit ont été étudiés sur plus de 2 000 génotypes interconnectés via leurs pedigrees. Les héritabilités et corrélations génétiques entre caractères ainsi qu'entre valeurs mesurées ou estimées par un panel de dégustateurs ont été finement analysées (Kouassi et al., 2009).
De nombreux QTL(1) ont été localisés pour chaque caractère étudié : 15 pour la texture de la chair, certains co-localisant avec des gènes intervenant dans la biosynthèse de l'éthylène (Md-ACO1, Md-ACS1 ; Costa et al., 2005), d'autres correspondant à des enzymes de dégradation des parois cellulaires (Mathis et al., 2008).
Les composés aromatiques ont été récemment corrélés avec des clusters(2) de gènes (Dunemann et al., 2009). Les corrélations génétiques existant entre ces composés commencent à être décryptés (Rowan et al., 2009).
Enfin des caractères plus « classiques » type coloration pourraient être revisités dans les programmes de sélection suite à la révélation de leur contrôle génétique de la couleur rouge (Chagné et al., 2007).
Variabilité des résistances
Concernant la résistance aux maladies et parasites, l'ampleur de la variabilité génétique naturelle varie selon le bio-agresseur considéré.
Elle est très forte pour la résistance à des maladies comme la tavelure ou l'oïdium pour lesquelles des gènes majeurs de résistance et des sources de résistance partielle ont été simultanément décrits (pour la tavelure, voir Gessler et al., 2006 ; pour la résistance à une large gamme de pathogènes et bio-agresseurs, voir Alston et al., 2000).
Les principaux gènes majeurs ont été identifiés chez des espèces sauvages de Malus (ex. : Malus floribunda et Malus micromalus comme origine respective des gènes de résistance à la tavelure Vf et Vm, ou Malus robusta et Malus zumi comme origine respective des gènes de résistance à l'oïdium Pl1 et Pl2). Mais des gènes majeurs ont aussi été caractérisés chez le pommier domestique (ex. : Vg ou Vd chez les variétés Golden Delicious ou Durello di Forli respectivement Bénaouf et Parisi, 2000 ; Tartarini et al., 2004).
Les résistances partielles ont été recherchées surtout chez le pommier domestique (Schmidt., 2000 pour la tavelure ou Visser et al., 1974 pour l'oïdium) et cartographiées à partir de certaines variétés (Calenge et al., 2004 ou Soufflet-Freslon et al., 2008, pour la tavelure ; Calenge et Durel, 2006 pour l'oïdium).
Le type et l'origine des facteurs de résistance semblent similaires vis-à-vis du feu bactérien (Peil et al., 2007 ; Durel et al., 2009 ; Calenge et al., 2005).
Pour le chancre à Nectria, des résistances partielles sont décrites (Lateur et Populer, 1994 ; Sasnauskas et al., 2006) mais la variabilité semble importante. Pour d'autres maladies secondaires (ex. Phyllosticta solitaria), la variabilité génétique de la résistance a été moins explorée ou moins décrite.
C'est aussi le cas pour certains insectes chez lesquels il est plus difficile d'évaluer un niveau de résistance. Cependant, une forte variabilité a été décrite pour la résistance au puceron cendré Dysaphis plantaginea (Rat-Morris, 1994 ; Arnaoudov et Kutinkova, 2006 ; Miñarro et Dapena, 2007). La cartographie de QTL a même été tentée pour le puceron cendré et le puceron à galles rouges D. devecta (Stoeckli et al., 2008). En revanche, la variabilité génétique semble très faible pour la résistance au carpocapse Cydia pomonella.
Soulignons que, pour estimer l'ampleur de la variabilité de la résistance aux bio-agresseurs, il peut être important de considérer la corrélation entre les symptômes observés sur feuilles et sur fruits si les deux types d'organe sont l'objet d'attaques. De même, le niveau de résistance peut évoluer avec l'âge des plantes. C'est le cas de l'oïdium (corrélation juvénile-adulte faible), ou de la relation entre le niveau de pression parasitaire et l'environnement (interaction génotype x milieu potentiellement exploitable en sélection).
Variabilité architecturale
Bernhard (1961) puis Lespinasse (1977) ont observé une large variabilité du port des arbres chez le pommier. Ils ont classé les variétés selon quatre types, classification légèrement revue par Lespinasse en 1992. Cela va des variétés de type I avec leur port colonnaire typique (dû à la mutation du gène Co, ex. Wijcick ; Tobutt, 1994) jusqu'aux variétés de type IV à port retombant et ramification principalement acrotone avec un angle de branchement ouvert et une fructification à l'extrémité des rameaux longs et moyens (ex. Granny Smith, Fuji).
Mais cette typologie ne suffit pas à caractériser finement le comportement de croissance, ramification et fructification des variétés. On sait que des variétés type Fuji, bien que classées en type IV, peuvent avoir une production très alternante entre années consécutives.
Depuis le début des années 1990, un ensemble d'études a montré que le caractère de régularité de production pouvait correspondre à deux stratégies (Lauri et al., 1995, 1997 ; Lauri et Laurens, 2005). Dans la première, la production est régulière sur chaque rameau fructifère individuel. La seconde, plus complexe, correspond à une désynchronisation des rameaux fructifères : chaque année une partie des rameaux est à fruit et l'autre végétative. Cette désynchronisation entre branches d'un même arbre voire, au sein d'une même branche, entre rameaux portés par des « âges de bois » différents est souvent observée.
Les bases physiologiques sous-jacentes à ces phénomènes commencent à être élucidées. La longueur de l'axe pourrait jouer un rôle primordial dans la fréquence de floraison terminale, probablement via la surface foliaire (Lespinasse, 1970 ; Lauri et Trottier, 2004). Mais d'autres phénomènes tels que la densité de ramification interviennent. Ainsi, les variétés régulières en production présentent souvent un avortement important des ramifications florifères à un stade précoce. C'est le phénomène d'extinction.
Depuis cinq ans environ, des études sont menées sur le déterminisme génétique des caractères architecturaux, en analysant une descendance en ségrégation issue du croisement de deux parents à port contrasté, Starkrimson et Granny Smith. De nombreux caractères de croissance (nombre d'entre-nœuds par pousse, longueurs des pousses), de ramification (nombre et répartition des rameaux axillaires par catégories de longueur), de géométrie (angles de branchement, volume de l'arbre) se sont révélés héritables au cours de la phase juvénile des hybrides (Segura et al., 2006).
Plusieurs QTL ont été détectés (Segura et al., 2007 ; 2008) et leurs effets décomposés selon l'année climatique et l'âge de l'arbre (Segura et al., 2009). La classification de ces hybrides sur la base de ces caractères a montré que seuls quelques-uns pouvaient être rapprochés des types I à IV alors qu'une grande majorité présentait des caractères recombinés. Par exemple, certains ont des ramifications plutôt courtes mais des diamètres faibles et des branches plutôt ouvertes (Segura et al., 2009b, figure 1). Ces recombinaisons de caractères sont en cours d'étude durant la phase mature des arbres et en considérant d'autres croisements.
Variabilité des caractères adaptatifs
Très récemment, des études ont été initiées sur les caractères d'adaptation aux conditions environnementales : recherche d'optimisation entre économie d'eau (par régulation stomatique) et acquisition du carbone (pour maintenir le calibre de fruit et la régularité de production).
En effet, en conditions chaudes, l'arbre augmente sa transpiration tant que l'eau est disponible dans le sol. Mais l'irrigation pourrait être limitée à l'avenir, d'où des situations de stress hydrique plus ou moins longues et/ou sévères. De tels stress provoquent la fermeture des stomates pour diminuer la perte en eau par transpiration, d'où un échauffement de la surface de la feuille et une réduction de l'acquisition du carbone nécessaire au développement du fruit et de l'arbre.
Des stratégies différentes d'utilisation de l'eau ont été mises en évidence entre les variétés Fuji et Braeburn (Massonnet et al., 2007). Braeburn est plus économe que Fuji par limitation stomatique des échanges gazeux.
Sur la base de ces résultats, la descendance Starkrimson x Granny Smith a été explorée pour la réponse de l'arbre à des conditions de stress abiotique, en conditions contrôlées et en verger. Les réponses foliaires au déficit de pression de vapeur de l'air sont fortement héritables et quatre QTL ont été détectés : trois relatifs à la conductance stomatique et à la transpiration, et un au taux de photosynthèse. La relative indépendance des 2 QTL majeurs ainsi que les effets alléliques ont permis d'identifier des hybrides recombinants à bonne efficience d'utilisation de l'eau (Regnard et al., 2009).
Des recherches sont en cours sur la réponse foliaire au stress hydrique estival, et d'autres caractères doivent encore être explorés : manque de froid en période automno-hivernale, effet de températures élevées.
Construction d'idéotypes variétaux
Des enjeux complexes
La construction d'idéotypes variétaux s'inscrit dans un cortège de priorités impliquant divers acteurs (producteurs, transformateurs, metteurs en marché, consommateurs, instituts techniques et de recherche) aux objectifs divers.
Les enjeux économiques sont cruciaux pour les producteurs et metteurs en marché et les enjeux environnementaux et de santé sont portés en priorité par les consommateurs. Mais des convergences sont possibles.
Ainsi les producteurs ont intérêt économiquement et pour leur santé à réduire les traitements.
De Buck et Buurma (2004) proposent de représenter ces objectifs contradictoires sous forme d'un « triangle des trois P » : People, Planet, Profit. Ils soulignent que l'émergence d'un consensus faciliterait l'organisation des recherches et des politiques publiques nationales, européennes voire internationales.
Concernant l'interaction entre plantes et bio-agresseurs, la construction d'idéotypes variétaux adaptés aux bas intrants amène une rupture avec la façon de penser la sélection de pommiers résistants qui a prévalu durant les trois dernières décennies. Le terme « construction » est à prendre au sens large : il ne s'agit pas seulement de construire des génotypes mais de connaître les méthodes culturales permettant de les valoriser dans le contexte pédoclimatique et le système de culture les mieux adaptés à leurs potentialités.
Interactions plante-bioagresseurs : le cas de la tavelure
Un exemple parlant est celui de la résistance à la tavelure, principale maladie fongique du pommier. Les programmes d'amélioration variétale ont longtemps privilégié la résistance monogénique. C'est la plus facile à sélectionner (test fiable au stade plantule), puis à utiliser par l'arboriculteur (pas de traitement... au début). Ce progrès génétique réalisé aurait permis une réduction d'environ 75 % des traitements fongicides (Audemard et al., 1995) si les variétés avaient eu du succès commercial et si la résistance avait été stable. Or elles sont encore peu cultivées (1 à 2 % de la surface de pommiers actuellement) et la résistance conférée par le gène Vf a déjà été contournée en Europe (Parisi et al., 1993, 2004). Des traitements visant à retarder l'apparition et la propagation des souches virulentes ont été préconisés. Ceci limite la réduction des fongicides : 30 à 61 % de réduction suivant les bassins de production (Sauphanor et al., 2009).
Certes, les variétés récentes à résistance monogénique de type Ariane se comportent bien en verger commercial et fournissent une production de qualité en utilisant moins de fongicides. Mais certaines de ces variétés, une fois contournées, pourraient être très sensibles à la maladie : la résistance monogénique peut avoir été introduite sur un fonds génétique dépourvu de résistance quantitative (cette dernière assure à la variété sa « rusticité » ou capacité à ralentir les épidémies et à en limiter les dégâts). La réduction de l'indice de fréquence de traitements (IFT) risque donc de n'être pas durable.
Face à Venturia inaequalis, champignon à reproduction sexuée obligatoire tous les ans (source de variabilité), la meilleure solution en termes de durabilité semble bien la résistance polygénique (MacDonald et Linde, 2002). Celle-ci, plus difficile à sélectionner et à gérer car partielle et plus ou moins efficace suivant les années (selon les conditions climatiques et l'inoculum présent), doit être accompagnée de pratiques culturales et de traitements allégés. Sur Reine des Reinettes, variété à bon niveau de résistance partielle, nous avons montré que la réduction de traitements fongicides peut être de 47 à 54 % de l'IFT (par rapport à la moyenne régionale) sans prise de risque (Didelot et al., in press). Mais la mise en œuvre pratique des associations de méthodes à effets partiels demande beaucoup de technicité, ce qui peut freiner son adoption (Brun et al., 2007).
« Bas intrants », viser TOUS les bio-agresseurs à la fois
Un idéotype pour un verger à bas intrants ne vise pas la résistance à un seul bio-agresseur mais un bon comportement global au verger vis-à-vis d'un complexe de bio-agresseurs.
De plus, il n'y a pas « un » mais « des » idéotypes, en fonction des régions et du mode de production. Ils doivent être adaptés aux principaux bio-agresseurs présents et fortement dommageables dans chaque bassin de production. Dans certains cas comme en agriculture biologique, l'amélioration génétique doit viser en priorité certains bio-agresseurs (ex. puceron cendré) vu le manque de méthodes alternatives à la résistance variétale et compatibles avec le cahier des charges AB.
Par ailleurs, ces idéotypes doivent être adaptés aux conduites architecturales.
Ainsi, la conduite centrifuge (Lauri, 2002) réduirait les épidémies de tavelure. Pourquoi ? Des travaux ont montré que cette conduite augmente l'interception de lumière donc la précocité et la coloration des pommes (Willaume et al., 2004, figure 2). Nous en tirons l'hypothèse qu'elle réduit les durées d'humectation foliaire (Simon et al., 2006 ; thèse en cours à l'UMR PIAF, Clermont-Ferrand).
Mais il semble aussi que l'augmentation de la porosité de la frondaison apportée par la conduite centrifuge favorise le carpocapse (Simon et al., 2007).
Ainsi, une conduite architecturale défavorable à un bio-agresseur peut en favoriser un autre.
Ici comme ailleurs, il faut acquérir des connaissances sur le développement de l'arbre, en épidémiologie, écophysiologie et génétique, sur des gammes de génotypes cultivées dans des environnements et selon des pratiques variés, pour optimiser les caractéristiques des idéotypes. Dans cette démarche, il faut viser l'aptitude de la variété à ne pas subir de trop forts dommages directs ou indirects suite à une attaque de bio-agresseur : une forte sensibilité à la tavelure peut entraîner, par exemple, une chute précoce des feuilles affectant la mise à fruit de l'année suivante donc la régularité du rendement.
L'objectif n'est plus d'avoir une variété très productive en système intensif, mais une variété à production éventuellement moyenne mais régulière et de bonne qualité, capable de surmonter des conditions climatiques ou parasitaires difficiles.
Quelles stratégies pour l'amélioration ?
L'amélioration génétique du pommier conduite jusqu'à présent repose sur une variabilité génétique étroite et utilise surtout des génotypes « élites », type Gala, Braeburn ou Fuji (Noiton et Alspach, 1996). La variabilité des variétés locales a été insuffisamment exploitée. La raison majeure est qu'à chaque fois que l'on introgresse les caractères désirés (résistance, qualité gustative...) on en introduit aussi des rédhibitoires (alternance, conservation limitée...) Exploiter des variétés locales exige donc plusieurs cycles de croisements et sélection.
Au niveau des caractères eux-mêmes, des démarches méthodologiques récentes, développées dans le cadre du projet européen HiDRAS, ont révélé de nouveaux hybrides à forte valeur génétique et semblant intéressants en tant que géniteurs pour les caractères gustatifs du fruit (Kouassi et al., 2009).
Concernant les résistances, la stratégie actuelle vise à un pyramidage de gènes à effet partiel confortant l'action d'un ou quelques gènes majeurs. Une construction de ce type (gène majeur + QTL) s'est montrée efficace et difficile à contourner chez le piment (résistance aux potyvirus, Palloix et al., 2009). Pour la résistance du pommier à ses principaux agresseurs (tavelure, oïdium, puceron cendré...), la durabilité de cette stratégie reste à démontrer. On pourrait l'articuler avec des pratiques culturales (prophylaxie, traitements modérés, conduite de l'arbre...)
Les caractères de régularité de production et d'adaptation à des conditions de stress environnementaux restent encore peu considérés dans la sélection. On cherche encore des connaissances sur les méthodes de phénotypage adaptées au screening de larges populations et sur les déterminismes génétiques. La prise en compte de ces caractères reste donc à construire.
Au niveau des schémas de sélection, un effort a déjà été fait pour mieux articuler la recherche avec les mécanismes de sélection et de diffusion du matériel végétal. Par exemple, le partenariat INRA-Sarl Novadi (qui regroupe la plupart des pépiniéristes pommier) a conduit à la création de la SAS Pomalia qui a permis le succès de la variété Ariane (Laurens et Pitiot, 2002). D'autres types de collaborations pourraient se développer. Ainsi des réseaux participatifs (comme pour le blé dur, Desclaux et al., 2006) permettraient de disposer d'essais avec des niveaux d'intrants très variables et, ainsi, d'élargir les possibilités de sélection pour des gammes de conditions et modes de cultures allant de la haute densité en conduite raisonnée à l'agriculture biologique.
Un projet CTPS en cours géré par le Grab (Groupement de recherche en agriculture biologique) avec des partenaires INRA vise ainsi à mettre en place une méthodologie pour un réseau de sélection variétale décentralisé (Warlop et al., 2009).
L'apport de la modélisation...
Vu la nature multi-critères des idéotypes à concevoir et les évolutions à venir des conditions climatiques et des pratiques culturales, il faut explorer de nouvelles voies méthodologiques. Elles font appel à la modélisation des interactions génotype x environnement x conduite (IGEC).
Plusieurs types de modèles en cours d'essai utilisent des connaissances à diverses échelles de temps et d'espaces. Il y a trois niveaux possibles de modélisation :
– celle des interactions entre développement de l'arbre et cycle biologique d'un pathogène sur une année de croissance ;
– celle d'une parcelle tenant compte de l'espacement des arbres sur le rang, des rangs entre eux et de la composition variétale (mélange éventuel) ;
– celle du territoire avec plusieurs parcelles aux variétés et conduites variées.
... À l'échelle de l'arbre...
Des modèles de développement des arbres existent depuis la fin des années 70 et leur réalisme biologique a progressivement augmenté. Des modèles sont disponibles sur pêcher (Lopez et al., 2008 ; Génard et al., 2008) et pommier (Costes et al., 2008). Ils diffèrent par les processus représentés (allocation de carbone, répartition de la ramification, flexion des branches, etc.), et par les échelles de représentation du végétal. Des améliorations sont encore à effectuer.
Tout d'abord, représenter la relation entre phénologie de l'arbre et conditions climatiques permettrait de simuler, sur des scénarios climatiques probables, les interactions entre développement de la plante et des pathogènes, donc explorer les conditions permettant à l'arbre d'échapper à la(aux) maladie(s).
Autre amélioration : le couplage, en partie réalisé (Da Silva et al., 2008), des modèles de développement avec ceux d'interception de la lumière. Il permettra de calculer le micro-climat intra-arbre plus ou moins favorable aux pathogènes.
Enfin, il faut tenir compte de la variabilité génétique (présence/absence d'allèles et effets induits sur le phénotype). Les exemples existant sur la qualité de la pêche (Quilot et al., 2005) sont à transposer. Vaste chantier ! Ce défi majeur doit mobiliser des compétences à l'INRA et chez ses partenaires, pour prioriser les caractères à améliorer, désigner de nouveaux idéotypes et amorcer la boucle vertueuse modélisation-expérimentation (Figure 3).
... À plus large échelle
À une échelle plus large, des modèles représentant la parcelle et son dispositif de plantation permettraient d'estimer la propagation des pathogènes à moyenne distance (entre organes et entre individus). Un projet ARC INRIA-INRA est en cours sur la modélisation des échelles arbre-pathogène et parcelle sur le couple pommier-tavelure (http://www.sm.u-bordeaux2.fr/ M2A3PC/index.php). Toujours à l'échelle de la parcelle, des modèles de génétique des populations ont pour but de représenter l'évolution spatiale et temporelle de la plante hôte et du pathogène (Sapoukhina et al., 2009).
Enfin, aux échelles multi-parcelles et bassin de production, un modèle intègre l'effet des interventions culturales et des conditions pédoclimatiques voire économiques et sociales sur le phoma du colza. Il utilise le logiciel SIPPOMWOSR (collaboration INRA Grignon, INRA Rennes et CETIOM (Lô-Pelzer, 2008 ; Lô-Pelzer et al., 2008 ; Boillot et al., 2008).
Construction dynamique
Viser des cibles invariantes...
En conclusion, signalons un enjeu important : l'élaboration d'un idéotype de pommier aujourd'hui aboutira à une création variétale dans 15 à 20 ans au mieux. La nouvelle variété pourrait se révéler inadéquate lors de son inscription, sa diffusion ou son lancement si la réflexion n'intègre pas une vision prospective. La solution pourrait être d'évaluer la robustesse des idéotypes face à divers scénarios éco-socioenvironnementaux afin de définir des caractères cibles invariants, ou en tout cas le moins mouvants possibles.
La régularité de la production est ainsi sûrement un caractère à rechercher.
La résistance génétique est également souhaitable, sauf dans le cas où son acquisition défavorise un autre caractère majeur type qualité (à cause d'un antagonisme métabolique par exemple). Attention aussi à la résistance totale monogénique vis-à-vis d'un seul pathogène (ex. tavelure) : elle risque de ne pas être durable et de favoriser d'autres maladies suite à la baisse des traitements.
... Et savoir évoluer ensuite !
En fait, la construction d'un idéotype à un moment donné intègre l'ensemble des informations disponibles à cet instant-là. Mais c'est un processus dynamique appelé à évoluer ensuite en fonction de l'acquisition de nouvelles connaissances.
Il est raisonnable de penser que des premières cibles seront définies en fonction du contexte actuel, puis progressivement validées ou élargies par l'exploration de modèles notamment écosocio-environnementaux. Ainsi, l'approche multi-critères basée sur des modèles qualitatifs fournit des bilans sur la performance, les risques ou la durabilité des systèmes culturaux (Znidarsic et al., 2006). L'INRA Grignon développe une telle démarche dans le cadre du réseau européen Endure (http://www.endure-network. eu/), à l'aide du logiciel DEXi (Bohanec et al., 2008), pour réaliser une approche multi-critères de systèmes de culture innovants (exemple pour la vigne : Delmotte et al., 2008).
Les idéotypes ainsi conçus seront confrontés à la variabilité génétique et aux processus d'innovation et de sélection dans lesquels les réseaux participatifs auront probablement une place plus grande pour répondre aux attentes des divers systèmes de culture.
On pourra ainsi espérer un cercle vertueux entre expérimentation, modélisation/prédiction et innovation, impliquant un grand nombre d'acteurs et de compétences.
<p>* UMR 1098 Développement et Amélioration des Plantes (DAP), Équipe Architecture et Fonctionnement des espèces fruitières (AFEF), CIRAD, Avenue Agropolis, Montpellier.</p> <p>** UMR Génétique et Horticulture (GenHort), INRA-Agro-CampusOuest-UA, Centre INRA Angers-Nantes, BP 60057 49071 Beaucouzé Cedex.</p> <p>*** INRA UERI de Gotheron, Domaine de Gotheron 26320 Saint-Marcel-lès-Valence.</p>
1 - Qu'est-ce qu'un « idéotype » ?
La notion d'idéotype, introduite par Donald (1968), consiste à définir explicitement les plantes « souhaitées/désirées » pour un environnement et un type d'agriculture donnés, sur la base de la variation des caractères cibles et de leurs interactions.
Cette notion a été utilisée sur espèces annuelles (e.g. Yin et al., 2003) et pérennes forestières (Wu, 1998) et fruitières (e.g. Dickman et al., 1994). Sur ces dernières, des propositions ont été faites à propos de l'interaction entre développement de l'arbre et production (Lauri et Costes, 2004 ; Cilas et al., 2006).
Les caractères cibles sont très divers. Ils concernent en priorité la qualité des produits et leur capacité de stockage post-récolte, mais aussi la résistance aux maladies, l'adaptation au stress hydrique, la facilité de conduite de l'arbre et sa régularité de production. Or, chaque caractère pris séparément est le plus souvent contrôlé par un déterminisme génétique complexe impliquant de nombreux gènes à effets faibles. La conception d'un idéotype doit donc s'appuyer sur des combinaisons judicieuses d'allèles plutôt que sur des gènes majeurs seuls. De plus, la plupart des caractères, comme la résistance aux bio-agresseurs ou le développement de l'arbre, sont influencés par des facteurs non génétiques : environnement pédo-climatique, populations de pathogènes ou manipulations appliquées à l'arbre.
Enfin, la conception d'idéotypes revêt une complexité supplémentaire chez les arbres fruitiers. En effet, l'expression des caractères peut évoluer en fonction de l'âge des arbres (effet d'ontogénie) et être affectée par des effets cumulatifs entre années consécutives. Par exemple l'effet d'un stress hydrique peut réduire la croissance de l'arbre donc pénaliser la production de l'année suivante.
2 - Changement climatique : il met la pression

Tavelure sur fruits. Comme d'autres bio-agresseurs, elle peut être influencée par le changement climatique.
Sur le pommier, plusieurs effets du changement climatique sont déjà perceptibles. Les températures printanières et estivales plus élevées favorisent le nombre de générations de certains bio-agresseurs. Ainsi on observe maintenant trois générations par an du carpocapse dans le Sud-Est de la France.
Depuis la fin des années 80, les dates de floraison moyennes ont avancé. Cela résulte de deux phénomènes contradictoires : une satisfaction des besoins en froid en période automno-hivernale plus lente et une arrivée plus précoce des températures chaudes en début de printemps (Legave et al., 2008). Outre des problèmes de pollinisation, ces changements modifient la phénologie de l'arbre et l'épidémiologie des bio-agresseurs très dépendants du microclimat de l'organe infecté (carpocapse, tavelure...)
Enfin, l'élévation des températures estivales augmente les besoins en eau de l'arbre et peut conduire à des stress hydriques voire thermiques. La production risque d'être pénalisée, ainsi que l'induction florale pour l'année suivante.
Figure 1 -

Hybrides de pommier ayant (A) un port retombant et une ramification surtout par rameaux courts ; les rameaux longs ont des diamètres faibles ; (B) un port intermédiaire : ramification peu abondante mélangeant rameaux longs et courts à forts diamètres. (D'après Segura et al., 2009b).
Figure 2 -
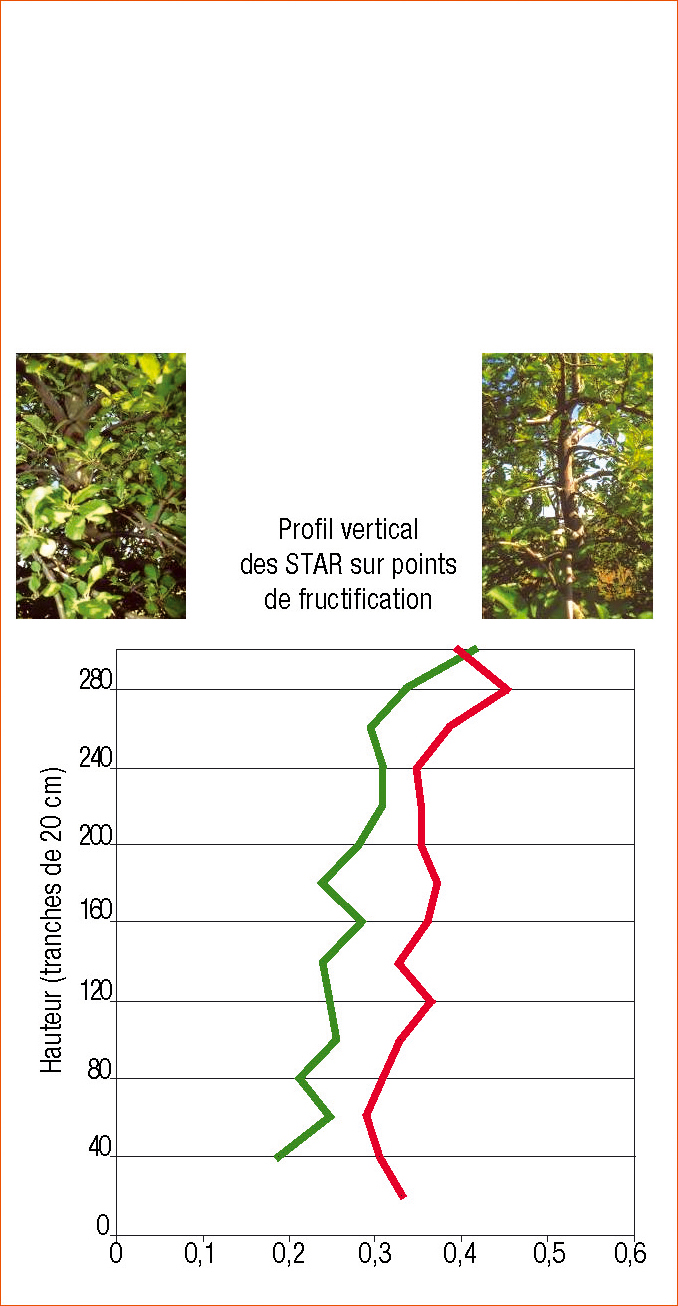
Efficience de deux systèmes de conduite (vert : système classique de type Solaxe; rouge : système de Conduite Centrifuge) sur l'interception de la lumière par le feuillage proche des points de fructification. STAR : Silhouette to Total Area Ratio. (D'après Willaume et al., 2004)
Figure 3 - La « boucle vertueuse » :
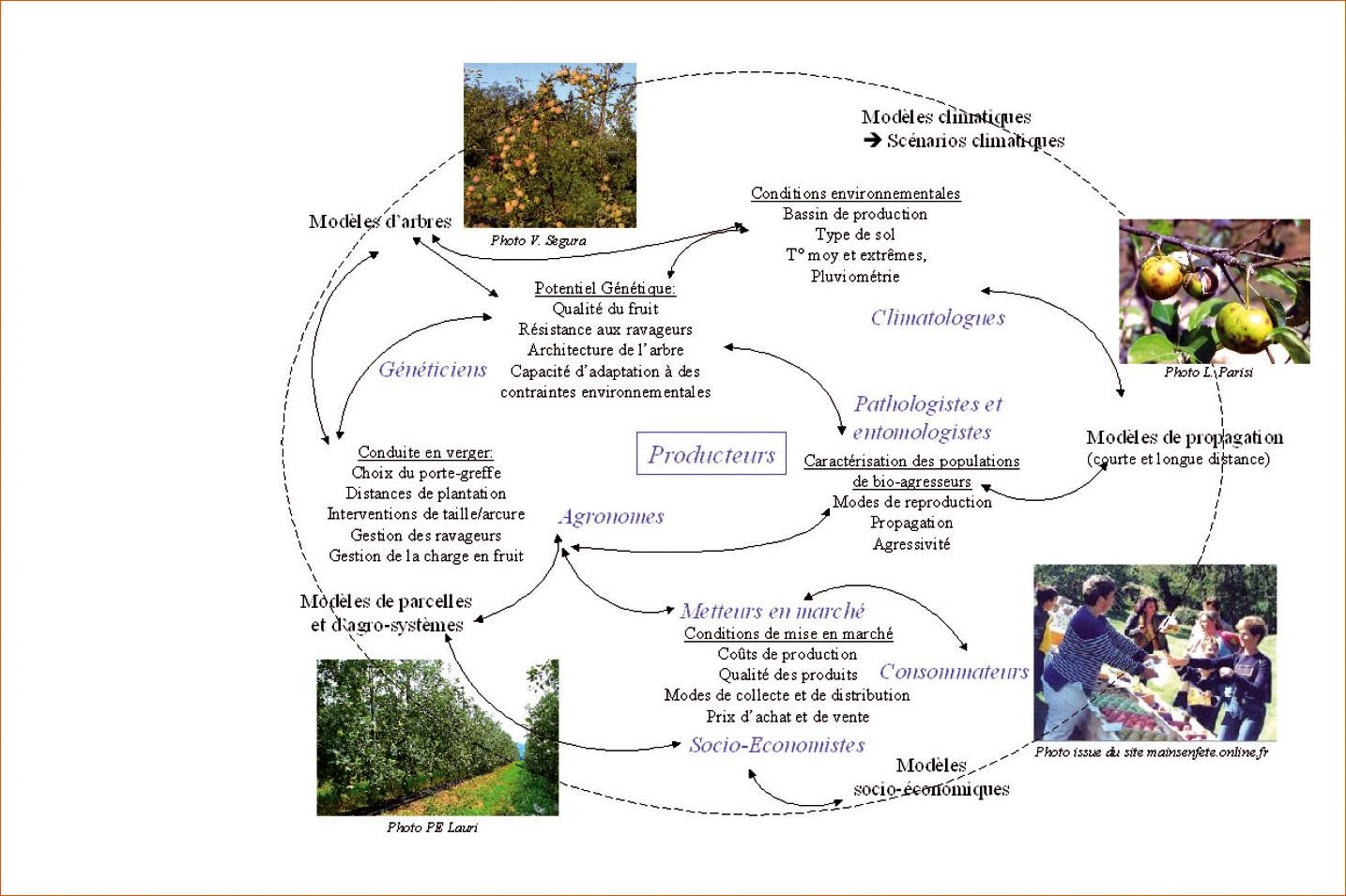
Schéma illustrant les différents acteurs (en bleu), les différentes disciplines et approches à fédérer pour concevoir, tester et rendre opérationnel un idéotype variétal de pommier.
3 - Situer cet article trouver sa bibliographie
Cet article s'inspire de la réflexion initiée par le groupe « Conception et évaluation d'idéotypes de pommier adaptés à des systèmes de culture économes en intrants », soutenu par l'INRA (DS Econat) dans le cadre de l'appel d'offres Ecophyto R&D : idéotypes pour des plantes cultivées en agriculture durable (groupe animé par Luciana Parisi).
Il cite une communication au CIAg (Carrefour de l'Innovation agronomique) du 5 novembre 2009 : « Variétés innovantes et modes de culture adaptés ».
Soulignons que cette communication est un rapport d'étape sur une réflexion à poursuivre et à étayer, d'autant qu'elle va conditionner des programmes de recherche à long terme.
Le texte intégral avec sa bibliographie (81 références) est disponible auprès des auteurs et sur : http://www. inra.fr/ciag/revue_innovations_agronomiques/volume_ 7_novembre_2009.

