
 Imprimer
Imprimer sur un morceau de carton ondulé contenant des larves de Cydia pomonella. X. Fauvergue - Inra")
Adulte femelle de Mastrus ridens (ovipositeur visible à l'arrière du corps) sur un morceau de carton ondulé contenant des larves de Cydia pomonella. X. Fauvergue - Inra

Spécimen femelle de M. ridens en collection. D. Muru - Inra
Le pommier et son principal ravageur le carpocapse Cydia pomonella, tous deux présents en France, viennent d'Asie centrale. Un organisme originaire de cette région du monde pourrait-il combattre « le carpo » ? Il existe un candidat pour cela. Son nom : Mastrus ridens.
Le carpocapse de la pomme
Un enjeu agronomique fort
La gestion des bioagresseurs en cultures fruitières ou légumières fait généralement appel à un ensemble de méthodes utilisées de manière coordonnée, qui peuvent être soit « génériques » (c'est-à-dire susceptibles de toucher simultanément plusieurs ravageurs), soit « spécifiques » d'un ravageur donné.
Malgré les efforts ainsi déployés, certains ravageurs demeurent malgré tout des problèmes majeurs. C'est le cas du carpocapse de la pomme, Cydia pomonella (lépidoptère, Tortricidae), responsable à lui seul de 30 % à 40 % des traitements insecticides-acaricides sur pommiers dans l'ensemble du territoire français, et également capable de toucher sérieusement d'autres cultures fruitières (poire, noix, coings).
Avec un total d'environ 12 millions de tonnes produites, la pomme est la première culture fruitière d'Europe. La France se classe au troisième rang européen (1,2 à 1,5 million de tonnes par an) derrière la Pologne et l'Italie(1).
Une caractéristique importante de cette culture est son niveau élevé de traitements phytosanitaires qui la place au deuxième rang en termes de quantité de pesticides par hectare, et au premier rang pour le nombre de traitements (Butault et al., 2011).
Une préoccupation majeure en PACA
Ces traitements ont pour cibles deux grandes catégories de bioagresseurs : les maladies fongiques et les insectes. En région PACA, première région productrice de pomme en France (Agreste Primeur 2015), plus de 40 % des traitements insecticides visent spécifiquement le carpocapse de la pomme (voir Encadré 1 p. 40). Cela en fait un enjeu prioritaire pour la réduction de l'utilisation de produits phytosanitaires. Même si plusieurs méthodes alternatives à la lutte chimique ont déjà été développées (granulovirusine, confusion sexuelle et filets « Alt'Carpo »), l'amélioration du contrôle du ravageur, en particulier dans les zones moins protégées (agriculture biologique, jardins, pommiers sauvages, etc.), reste une préoccupation majeure. Ceci est d'autant plus d'actualité que certaines de ces méthodes montrent des limites d'efficacité (résistance à la carpovirusine par exemple) et que le service envisagé ici serait « gratuit ».
L'auxiliaire potentiel : Mastrus ridens
Un auxiliaire de lutte biologique déjà évalué ailleurs
Une piste prometteuse pour améliorer le contrôle durable du carpocapse en France repose sur le déploiement d'une méthode dite de lutte biologique classique (Borowiec et al., 2011).
Brièvement, cette stratégie se définit comme l'introduction délibérée dans l'environnement d'un auxiliaire d'origine exotique en vue de son établissement pérenne et d'un contrôle durable du ravageur ciblé. L'introduction récente de Torymus sinensis contre le cynips du châtaignier en est l'exemple même d'une opération aboutie (Borowiec et al., 2013).
Indices prometteurs en Californie
Le projet présenté ici vise à introduire et acclimater l'insecte parasitoïde(2) Mastrus ridens (hyménoptère, Ichneumonidae). Cet insecte est issu de l'aire d'origine du carpocapse (Asie Centrale) (Kuhlmann et Mills 1999). Il a déjà fait l'objet d'introductions comme auxiliaire en Californie (1995) et, plus récemment, en Argentine (2005), au Chili (2006), en Nouvelle-Zélande (2012) et en Australie (2015).
Il s'est établi avec succès dans toutes ces régions (Mills 2005 ; Tortosa et al., 2014 ; Sandanayaka unpublished data ; Hossain unpublished data ; Zaviedo unpublished data). Si le gain économique dans ces régions reste à l'heure actuelle difficile à évaluer, les résultats acquis en Californie montrent un établissement systématique avec des taux de parasitisme sur cocons hivernants de carpocapse supérieurs à 40 % et pouvant atteindre 70 %, ainsi qu'une réduction significative des populations de carpocapse dans certains vergers (Mills 2005). Autant d'indices prometteurs en vue de l'introduction de cet auxiliaire-candidat en France.
Une opération conforme avec la réglementation
Dossiers expertisés par l'Anses
Mastrus ridens étant une espèce exotique, son introduction sur le territoire français est soumise à l'arrêté AGRG1225395A. Celui-ci impose d'abord le dépôt d'une demande d'introduction en milieu confiné auprès du ministère chargé de l'Agriculture puis, en second temps, le dépôt d'une demande pour l'évaluation au champ de l'auxiliaire.
Dans les deux cas, l'Anses expertise les dossiers sur la base d'évaluations réalisées par des experts extérieurs et fournit son avis au ministère de l'Agriculture qui rend l'arbitrage final. Le dossier d'introduction en milieu confiné a été facilité par notre quarantaine dédiée, Entomopolis (Thaon et al., 2017).
Importation de spécimens
Notre demande ayant été validée le 19 mai 2015, un premier lot d'environ 1 000 M. ridens a été importé depuis la Nouvelle-Zélande (LOA FR15PA00001 du 22/05/2015). Ensuite, afin de disposer de souches d'origines différentes, nous avons également importé des spécimens provenant de l'université catholique du Chili (LOA FR16PA00001 du 11/01/2016 et LOA FR16PA00002 du 01/03/2016).
Concernant la demande d'évaluation au champ, l'objectif est de fournir aux évaluateurs le maximum d'informations afin d'objectiver la « pesée d'intérêts » entre les bénéfices attendus et les éventuels risques (effets non intentionnels). Pour cela, nous nous sommes évidemment appuyés sur les travaux antérieurs menés à l'occasion des introductions dans d'autres pays, mais, en outre, nous avons plus particulièrement contribué à améliorer les connaissances dans deux domaines : la caractérisation moléculaire des populations de M. ridens (Retamal et al., 2016) et l'étude de la spécificité d'hôte. Cette dernière caractéristique est majeure pour l'efficacité et l'innocuité de l'auxiliaire.
Évaluation de la spécificité de Mastrus ridens
Tests sur dix espèces non-cibles
À l'occasion de précédentes introductions, la spécificité d'hôte de M. ridens avait été documentée vis-à-vis de dix-sept espèces de lépidoptères. Ces études avaient conclu que l'introduction de M. ridens présentait un risque très faible pour les espèces non-cibles aux États-Unis, au Chili, en Argentine, Nouvelle-Zélande et Australie (Charles et al., 2013).
Afin de compléter les données sur la spécificité d'hôte du parasitoïde, nous avons donc testé la capacité d'oviposition de femelles de M. ridens vis-à-vis de différentes espèces de lépidoptères présents en France. Ces espèces ont été choisies en fonction de leur proximité phylogénétique (classification taxonomique) et/ou de leur similarité écologique avec le carpocapse (développement sur les mêmes plantes-hôtes par exemple). Au total, nous avons testé la spécificité de l'auxiliaire vis-à-vis de dix espèces non-cibles (identifiée en gras dans le tableau ci-contre). Ce sont, des plus proches au plus éloignées phylogénétiquement :
- des espèces appartenant au genre Cydia (Cydia nigricana et Cydia splendana) ;
- des espèces d'autres genres appartenant à la même tribu (Grapholitini) que le carpocapse (Grapholita molesta, Pammene sp.) ;
- des espèces appartenant à d'autres tribus de la sous-famille des Olethreutinae (Lobesia botrana) ;
- des espèces appartenant à la sous-famille des Tortricinae (Eupoecilia ambiguella) ;
- en complément, des représentants de deux vastes familles de lépidoptères (« out-group ») susceptibles d'être rencontrés par M. ridens : Agrotis ipsilon, Mamestra brassicae (Noctuidae) et Ephestia kuehniella, Galleria mellonella (Pyralidae).
Résultat en conditions de « non-choix »
Pour chacune de ces espèces non-cibles, nous avons testé vingt femelles de M. ridens (accouplées et âgées de sept jours) en les isolant individuellement à l'intérieur d'une boîte de Petri et en les mettant en présence d'un seul cocon de l'espèce à tester (conditions de non-choix). Au bout de 24 h, chaque femelle de parasitoïde était retirée et le développement des chenilles était suivi jusqu'à émergence des adultes papillons ou parasitoïdes.
Les résultats obtenus (Figure 1) ont confirmé la spécificité de M. ridens. Dans ces conditions de non-choix et en espace très restreint, M. ridens ne s'est intéressé qu'à deux des dix espèces non-cibles testées. Ces dernières appartiennent au genre Cydia (C. nigricana et C. splendana) et présentent des résultats contrastés et toujours significativement différents de ceux observés sur C. pomonella (95 % de larves parasitées). Pour C. splendana, on observe 10 % des larves parasitées par M. ridens tandis que cette proportion est de 75 % pour C. nigricana. Chez cette dernière, on observe toutefois 15 % de larves mortes. Sachant que cette mortalité n'est pas constatée sur les lots témoins, cela suggère que ces larves ont été attaquées par M. ridens mais que le développement de ce dernier n'a pas pu se faire.
De la même manière, le nombre moyen de descendants par larve parasitée diffère selon l'espèce considérée. Il est toujours significativement plus important chez C. pomonella (moyenne de cinq descendants par larve) que chez C. nigricana (moyenne de deux descendants par larve) et C. splendana (moyenne de 1,5 descendant par larve).
Résultats en conditions de choix
En conditions de choix, chaque femelle de parasitoïde a été mise en présence de deux cocons de l'espèce à tester et deux cocons de C. pomonella. Dans ces conditions, deux lots de larves de C. nigricana sur cinq ont été parasités mais seulement lorsque les larves de C. pomonella ont été préalablement parasitées.
Ces résultats tendent à montrer que, en situation de choix et en conditions limitantes en hôtes, les larves non-cibles de C. nigricana ne sont que faiblement parasitées par M. ridens, et apparemment uniquement après exploitation de l'ensemble des larves de C. pomonella disponibles.
Une espèce spécifique du genre Cydia, avec une préférence nette pour C. pomonella
Au total, la spécificité de M. ridens a donc été testée vis-à-vis de vingt-cinq espèces de lépidoptères présentant des similarités phylogénétiques et écologiques variées par rapport à C. pomonella (voir tableau).
Ces tests montrent que :
- à ce jour, aucune espèce n'appartenant pas à la famille des Tortricidae n'a été parasitée par M. ridens en conditions contraintes (non-choix, espace restreint, conditions de laboratoire) ;
- au sein des Tortricidae, les tests effectués ailleurs dans le monde ont montré des ovipositions par M. ridens de quelques espèces n'appartenant pas au genre Cydia (Hennessey et al., 1995 ; Charles et al., 2013) ; cependant, dans chacun de ces cas, soit il n'y avait pas de parasitisme réussi (émergence d'adultes de parasitoïdes), soit le développement du parasitoïde était qualifié d'anormal sur la base du nombre, de la taille ou de la longévité des descendants ;
- seules quelques espèces appartenant toutes au genre Cydia (C. nigricana, C. splendana et C. succedana) ont fait l'objet d'un parasitisme réussi par M. ridens, mais avec un succès toujours plus faible que sur C. pomonella, que ce soit en nombre ou en taille de descendants.
En France, le genre Cydia est représenté en France par trente-deux espèces (De Jong et al., 2014), dont beaucoup se développent sur des plantes-hôtes particulières (par exemple C. nigricana est signalé sur des plantes appartenant à la famille des Fabaceae), ce qui rend a priori très faible leur probabilité de rencontre avec M. ridens.
Vers l'introduction et l'évaluation de Mastrus ridens au champ
Réseau de sites de lâchers
Sur la base des introductions antérieures et des éléments acquis par notre laboratoire, l'Anses et la DGAL ont délivré un avis favorable à l'introduction de M. ridens en France(3). Cela s'est traduit par la délivrance d'un arrêté des ministères de l'Agriculture et de l'Environnement (le 09/05/2017) autorisant les lâchers de M. ridens. Il convient donc désormais de se tourner vers l'évaluation au champ de M. ridens. Cela représente un défi en premier lieu financier. En effet, les financements acquis jusqu'à présent ne permettent que le soutien des activités de prélâchers, puis technique et logistique.
À l'image de ce qui a été fait lors d'opérations antérieures - tentatives d'introductions de Psyttalia lounsburyi contre la mouche de l'olive (Malausa et al., 2010) ou introductions réussies de T. sinensis contre le cynips du châtaignier (Borowiec et al., 2013) -, l'objectif est de mettre en place dans les meilleurs délais un réseau de sites de lâchers pertinent et cohérent. Nous estimons qu'il en faudra probablement plus de cinquante.
De telles opérations représentent le meilleur moyen pour évaluer rigoureusement et rapidement le potentiel de l'auxiliaire et adresser des questions de recherche (voir Encadré 3) permettant in fine l'optimisation des pratiques de recherche et développement en lutte biologique, pas seulement contre le carpocapse.
(1) Source : FranceAgriMer.(3) Lien à télécharger sur www.anses.fr/fr/system/files/MASTRUSRID_MO16-005_Ans.pdf.Lien de l'arrêté : https://info.agriculture.gouv.fr/gedei/site/bo-agri/document_administratif-43b1a58f-f360-4bac-8dd3-f5f0eeb142b6
Fig. 1 : Spectre d'hôtes de M. ridens au laboratoire
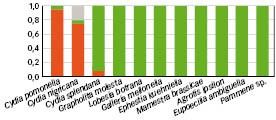
Larves « parasitées » (en rouge), « saines » (en vert) et « mortes » (en gris) chez différentes espèces de lépidoptères mises en présence de femelles de M. ridens en situation de non-choix (chaque femelle est mis en présence d'une seule larve de l'espèce à tester). La catégorie « mortes » concerne les hôtes retrouvés morts après une ou plusieurs ovipositions, mortalité induite par le venin injecté et/ou le développement incomplet de M. ridens.
1 - Éléments de biologie du carpocapse de la pomme et de Mastrus ridens

Cage d'élevage de M. ridens Photos : N. Borowiec - Inra

Rouleau de carton ondulé contenant des cocons de carpocapse. Photos : N. Borowiec - Inra
Les larves du carpocapse de la pomme se développent à l'intérieur des fruits où elles se nourrissent des pépins. Une fois parvenues au dernier stade de développement, elles sortent des fruits pour aller tisser un cocon (dans une anfractuosité de l'arbre ou dans le sol) où elles se nymphoseront.
Ce papillon possédant deux à trois générations annuelles, les larves des cocons de première et deuxième générations donneront principalement des adultes. Celles présentes dans les cocons de troisième génération entreront en diapause pour ne donner des adultes qu'au printemps suivant.
Mastrus ridens est un ectoparasitoïde grégaire, c'est-à-dire que l'adulte femelle pond ses oeufs sur la larve-hôte (et non pas à l'intérieur de la larve comme chez les endoparasitoïdes), et que cette dernière pourra ensuite donner naissance à plusieurs adultes parasitoïdes.
Cette espèce a longtemps été confondue avec Mastrus ridibundus jusqu'à ce qu'Horstmann (2009) décrive officiellement l'espèce ridens et donne des critères morphologiques permettant la distinction de ces deux espèces.
Dans son aire native, M. ridens est décrit comme spécifique de Cydia pomonella dont il est un ennemi naturel (Mills, 2007). Les femelles de M. ridens sont spécifiquement attirées par les cocons de Cydia pomonella (Jumean et al., 2005). Elles s'attaquent donc au stade prénymphal, qui est le stade diapausant chez le carpocapse.
De ce fait, pour les élevages de l'auxiliaire au laboratoire, ce sont des cocons de C. pomonella qui sont utilisés et présentés aux femelles parasitoïdes. Pour les obtenir, des larves de carpocapse de dernier stade sont placées dans des rouleaux de carton ondulé afin d'y tisser leur cocon (photos ci-contre).
Étant donné le statut réglementaire de l'auxiliaire (voir p. 38-39), ces élevages sont maintenus dans des pièces climatiques situées à l'intérieur du bâtiment de quarantaine « Entomopolis » (UMR ISA, Sophia Antipolis) (Thaon et al., 2017).
2 - Partenariat mis en place à l'occasion de l'évaluation de Mastrus ridens
La mise en place de ce projet a nécessité de nombreuses collaborations nationales et internationales. La première étape a été d'initier les élevages de M. ridens et de son hôte dans nos laboratoires, sur le site Inra de Sophia Antipolis. Sur ce point, nous avons bénéficié de l'expérience de collègues de l'Inra d'Avignon (UMR « Plantes et systèmes de cultures horticoles ») qui dispose d'élevages de carpocapse depuis plusieurs années.
Nous avons collaboré également avec la société Bioline AgroSciences (ex-Biotop) pour la fourniture de larves de carpocapse le temps que nos élevages de ravageurs soient opérationnels. L'obtention des souches et l'acquisition de la méthodologie d'élevage de M. ridens s'est faite en collaboration avec le « Plant and Food Research » en Nouvelle-Zélande.
La prochaine étape du projet consistera à évaluer au champ l'efficacité de M. ridens. La première évaluation, géographiquement « restreinte », sera effectuée en région PACA. Nous nous appuierons sur le réseau de vergers de la basse vallée de la Durance suivi par l'UMR PSH Inra (dispositif Sebiopag), pour lequel il existe un historique de suivi des populations de carpocapse et de parasitoïdes indigènes.
Des sites supplémentaires de PACA seront sélectionnés en lien avec la station expérimentale La Pugère et le GRCETA Basse Durance. Au total, l'évaluation au champ de M. ridens devrait se faire sur au moins cinquante sites. En cas de succès, l'étape suivante sera de déployer cette méthode de lutte biologique à d'autres bassins de production, voire d'autres cultures (poire, coing, noix).
3 - Installation de M. ridens : rôle des facteurs environnementaux et génétiques
Reposant sur l'introduction d'une espèce exotique, la lutte biologique classique peut être assimilée à une invasion biologique planifiée. Elle offre donc une opportunité unique d'étudier certains facteurs impliqués dans la réussite ou l'échec de l'établissement de ces populations, qu'ils soient démographiques (par exemple l'effet du nombre d'individus introduits ou du nombre d'introductions réalisées) ou génétiques (impact de la diversité génétique par exemple) (Fauvergue et al., 2012).
Ceci est d'autant plus important qu'environ deux tiers des opérations de lutte biologique classique se soldent par un échec de l'établissement des espèces introduites (Cock et al., 2016). Dans ce cadre académique, l'introduction de M. ridens en France permettrait d'aborder deux problématiques.
(1) Le rôle de la complexité de l'habitat sur l'établissement et la dispersion d'un auxiliaire de lutte biologique
La structuration spatiale des paysages agricoles est connue pour influencer la distribution et l'abondance des ravageurs et de leurs ennemis naturels. En revanche, peu de données sont disponibles concernant l'impact de cette structure sur la dynamique d'établissement et d'expansion d'un auxiliaire exotique. Il existe quelques évaluations théoriques et expérimentales réalisées en milieu contrôlé (Andrew and Ustin 2010, Hancock and Godfray 2012, Morel-Journel et al., 2015) mais il est difficile d'appliquer leurs résultats dans un environnement naturel, par définition beaucoup plus complexe. Dans le cadre des introductions planifiées de M. ridens en France, nous essaierons donc de mettre en évidence les facteurs démographiques, géographiques ou écologiques les plus déterminants lors de l'installation de l'auxiliaire.
Nous développerons également des modèles prédictifs permettant d'identifier les configurations de paysage favorables au contrôle biologique et de proposer des stratégies d'optimisation de déploiement des auxiliaires introduits.
(2) L'étude du vortex d'extinction chez un auxiliaire de lutte biologique
Chez certains hyménoptères, il existe un mode particulier de détermination du sexe (appelé CSD pour « Complementary Sex Determination ») qui provoque l'apparition de mâles non viables ou stériles (Zayed and Packer 2005).
Dans le cadre de petites populations introduites (cas de la lutte biologique classique), les faibles effectifs de départ peuvent entraîner une baisse de la diversité génétique qui elle-même augmentera la proportion de mâles non viables ou stériles, ce qui contribuera à diminuer les effectifs. Ainsi, le cycle se répète jusqu'à l'extinction possible de la population. C'est ce que l'on appelle le « diploid male vortex ».
Afin de tester ce phénomène, nous manipulerons la diversité génétique initiale de M. ridens via le nombre d'individus fondateurs puis nous utiliserons ces différentes populations pour les introductions expérimentales de terrain qui seront menées en France, mais également au Chili dans le cadre d'une collaboration avec l'université catholique du Chili.
Pour chacun de ces deux aspects académiques, il est certain que l'identification des leviers à l'établissement et à la dispersion de M. ridens permettra d'améliorer le potentiel de contrôle de l'auxiliaire en France, mais également en Europe.
REMERCIEMENTS Pour ce projet, l'Inra bénéficie du soutien financier de l'Onema dans le cadre de l'appel à projets PSPE2 du plan Écophyto (projet Bioccyd, 2015-2018, resp. : Nicolas Ris, Inra), du département Santé des plantes et environnement de l'Inra (projet Mastrus, 2016-2018, resp. : Xavier Fauvergue, Inra), et de l'Union européenne dans le cadre des appels à projets Marie Curie (projet Biomodics, 2013-2017, resp. : T. Malausa, Inra) et « ERA-net C-IPM » (projet API-Tree, 2017-2020, resp. : A. Alaphilippe, Inra).

