Imprimer
Imprimer
ph. M. Andrieux
Certaines résistances aux fongicides sont dues à des mutations de cible (Encadré p. 27). Elles sont dites croisées positives si le même champignon pathogène résiste à plusieurs substances différentes partageant le même mode d'action. Mais, parfois, des pathogènes résistent à plusieurs substances à modes d'action différents. Pourquoi ? Mutations empilées, chacune faisant résister à un fongicide ? Cela peut arriver (on parle alors de résistance multiple) mais il existe d'autres mécanismes. L'un d'eux aboutit aux résistances dites MDR, « multidrug resistance ». Le cas est connu en médecine humaine. Qu'en est-il des champignons phytopathogènes (1) en France ? Réponses dans cet article, tiré d'une communication de la session « Résistances aux fongicides » de la 9e CIMA (2).
La résistance MDR (se référer à l'encadré) des champignons pathogènes existe en pharmacie et en phytopharmacie. En pharmacie c'est-à-dire dans le domaine médical, elle s'observe en pratique chez divers pathogènes fongiques appartenant aux genres Candida et Aspergillus. Elle concerne particulièrement des triazoles de type IDM. En phytopharmacie – domaine agricole – de nombreux travaux menés par M. De Waard de l'Université de Wageningen aux Pays-Bas sur Botrytis cinerea et Mycosphaerella graminicola chez des mutants de laboratoire ont montré que la résistance MDR pouvait concerner les IDM mais aussi d'autres classes de fongicides (ex : phénylpyrroles, QoI).
Mais les investigations au champ n'ont démontré l'existence de ce phénomène que dans un nombre limité de cas. Trois d'entre eux ont été détectés en France chez B. cinerea, M. graminicola et Oculimacula yallundae, responsables respectivement de la pourriture grise de la vigne, de la septoriose du blé et du piétin-verse des céréales ; un cas concernant Penicillium italicum a été signalé au Japon.
L'objectif de cet article est de faire le point sur la résistance de type MDR rencontrée en France.
Matériel et méthodes
Souches fongiques isolées à partir de baies de raisin, de tiges et de feuilles de blé
Les souches de B. cinerea ont été collectées surtout dans le vignoble champenois au moment des vendanges, à partir de baies atteintes de pourriture grise. Elles sont maintenues sur un milieu à base de malt et d'extrait de levure, à 19 °C ; la sporulation est induite sous éclairage en lumière blanche.
Les souches d'O. yallundae isolées à partir de la base de tiges de blé et présentant des symptômes caractéristiques du piétin-verse sont issues d'essais conduits en France. Elles sont maintenues sur un milieu à base de farine de maïs, à 15 °C ; la sporulation est induite sous éclairage en lumière noire.
Les souches de M. graminicola isolées à partir de feuilles de blé portant des pycnides sont issues d'essais conduits en France. Elles sont maintenues sur un milieu à base de malt et d'extrait de levure, à 19 °C et à l'obscurité.
Pour P. yallundae et M. graminicola, les caractéristiques des divers phénotypes TriR résistants aux IDM sont précisés dans la littérature.
Essais sur du mycélium...
L'effet des fongicides sur la croissance mycélienne de B. cinerea et O. yallundae est étudiée après dépôt d'implants mycéliens calibrés sur un milieu de culture gélosé à base de glucose, sels minéraux et extrait de levure avec diverses concentrations des molécules testées.
Des mesures journalières (B. cinerea) ou hebdomadaires (O. yallundae) des diamètres des colonies mycéliennes permettent d'estimer la vitesse de croissance de chaque condition expérimentale puis la concentration inhibant de 50 % la croissance mycélienne (CI50 « mycélium »).
... et sur des spores
L'effet des fongicides sur l'élongation des filaments des trois champignons est étudié après dépôt d'une suspension de spores à la surface d'une eau gélosée (O. yallundae) ou d'un milieu gélosé à base de sucre et de phosphates (B. cinerea et M. graminicola).
Après incubation de 1 ou 2 jours (19 °C, obscurité), la longueur moyenne des tubes germinatifs est évaluée sous microscope à l'aide d'un micromètre oculaire. Pour chaque matière active, les essais conduits sur une gamme de concentrations permettent d'estimer celle inhibant de 50 % l'élongation des tubes germinatifs (CI50 « tubes germinatifs »).
Pour B. cinerea et M. graminicola, sur la base des essais sur spores, l'utilisation d'un milieu de culture à base de malt et d'extrait de levure et une incubation de 5 à 7 jours permettent d'évaluer les concentrations minimales inhibitrices (CMI) du développement fongique.
Pour tous les essais in vivo, les niveaux de résistance sont obtenus en calculant les rapports CI50 ou CMI « souches résistantes » / CI50 ou CMI « souches sensibles ».
Analyses de populations fongiques
Au niveau populationnel, des prélèvements de 15 à 30 organes végétaux malades par parcelle sont analysés après isolement (O. yallundae) ou non (B. cinerea, M. graminicola) des souches. Celles-ci sont caractérisées par des tests sur spores en utilisant des doses discriminantes de fongicides de modes d'action différents.
Résultats concernant la pourriture grise de la vigne
Vu depuis les années 1990
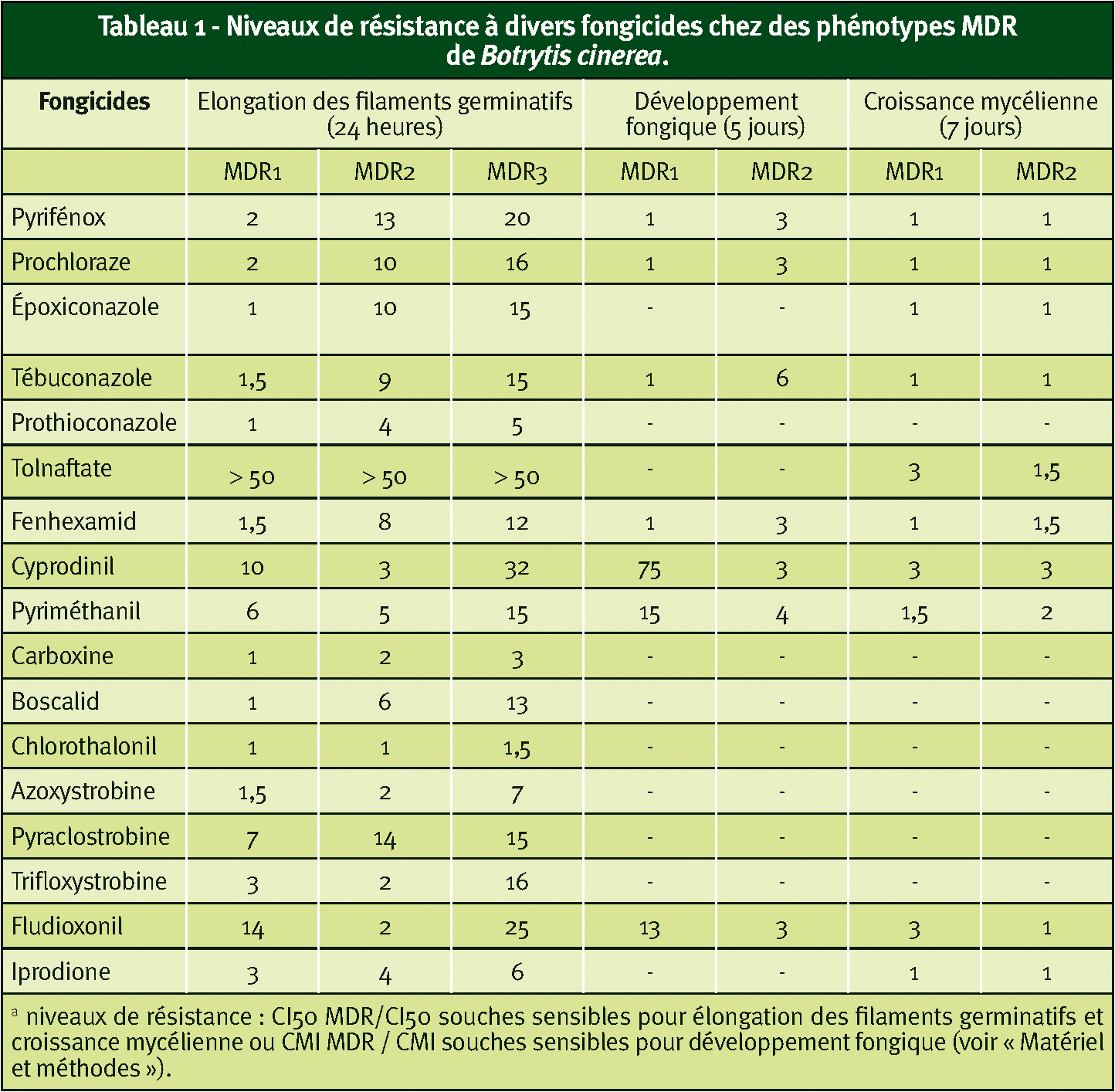
Lors de l'arrivée des anilinopyrimidines (ex : pyriméthanil, cyprodinil) et des phénylpyrroles (ex : fludioxonil) au début des années 1990, nous avons constaté la présence de souches de B. cinerea faiblement résistantes à ces deux anti-botrytis. Ce phénomène concerne en fait la plupart des fongicides utilisés contre la pourriture grise ainsi que des inhibiteurs stéroliques comme les IDM ou le tolnaftate (cette substance à usage médical affecte la squalene-epoxidase). Mais cette multi-résistance exprimée lors d'essais conduits sur spores l'est peu ou pas du niveau du mycélium (Tableau 1 p. 28).
Phénotypes MDR1 et MDR2 d'abord, MDR3 ensuite
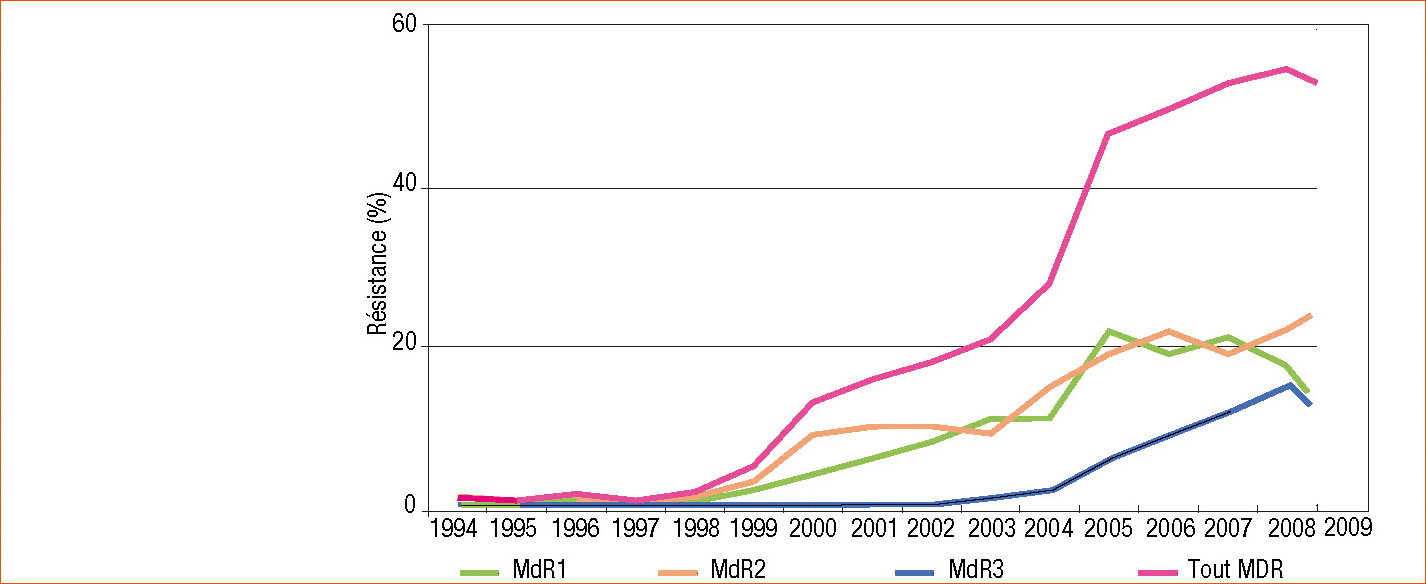
Dans un premier temps, nous avons, en fonction de leurs spectres de résistance croisée, identifié deux phénotypes : MDR1 (peu sensible au fluazinam et au fludioxonil) et MDR2 (peu sensible au fenhexamid, aux DMI et au boscalid). Pour chacun d'eux, la résistance est déterminée par un gène chromosomique. Les souches MDR3 plus tardivement détectées au vignoble et présentant le spectre de résistance le plus étendu sont des doubles mutants (Tableau 1).
Mécanismes caractérisés
Les recherches conduites en collaboration avec l'équipe de M. Hahn de l'Université de Kaiserslautern, en Allemagne, ont permis de caractériser le mécanisme moléculaire de cette multirésistance de type MDR.
Les souches MDR1 surexpriment l'ABC transporteur BcatrB, identifié antérieurement chez des mutants sélectionnés sur du fludioxonil. Ce phénomène résulterait de mutations dans un gène codant un facteur de transcription qui se lie au promoteur du gène BcatrB. Un mécanisme similaire existe en milieu médical chez des souches de C. albicans résistantes aux IDM.
Chez les souches MDR2, un transporteur à MFS est surexprimé suite à des changements dans le promoteur du gène codant ce transporteur.
Logiquement, les deux mécanismes sont impliqués pour les isolats multirésistants MDR3.
En pratique au vignoble
Cette mutirésistance de type MDR est décelée dans de nombreux vignobles européens mais elle est très fortement implantée en Champagne depuis quelques années. Elle concerne globalement la moitié des populations champenoises de B. cinerea avec actuellement une répartition à peu près équivalente des trois phénotypes (Figure 1 p. 28).
Ces souches MDR sont sélectionnées par les programmes anti-botrytis et il semble qu'elles présentent une fitness un peu moindre que les souches non MDR. L'impact pratique sur l'efficacité de la lutte chimique dans le vignoble champenois apparaît réduit.
Il faut enfin noter que la plupart de ces souches MDR ne présentent pas de résistance spécifique vis-à-vis des anti-botrytis récents (ex : anilinopyrimidines, fenhexamid, carboxamides).
Pour le piétin-verse des céréales
Depuis 1970, histoire mouvementée
La lutte chimique contre le piétin-verse a commencé dans les années 1970 avec l'utilisation des benzimidazoles et des thiophanates. Puis la généralisation de la résistance vis-à-vis de ces antimicrotubules a entraîné leur remplacement par des IDM du groupe des triazoles (ex : époxiconazole, flusilazole) ou des imidazoles (ex : prochloraze).
Des résistances ont également touché ces inhibiteurs stéroliques et le cyprodinil – une anilinopyrimidine affectant la biosynthèse d'acides aminés soufrés – a pu remplacer les IDM dans les régions particulièrement concernées.
Au début des années 2000, trois fongicides anti piétin-verse ont vu le jour : la métrafénone, le boscalid et le prothioconazole. Les deux dernières molécules ont retenu notre attention du fait de leur forte fongitoxicité in vitro.
Le boscalid est une carboxamide inhibitrice de la succinate deshydrogénase (SDHI) alors que le prothioconazole est un IDM proche des triazoles mais comportant un groupement triazolinethione. À noter que chez Oculimacula sp., cet inhibiteur stérolique ne présente pas de résistance croisée avec les triazoles ou le prochloraze.
Résistance MDR seulement chez O. yallundae, probablement TRiR1
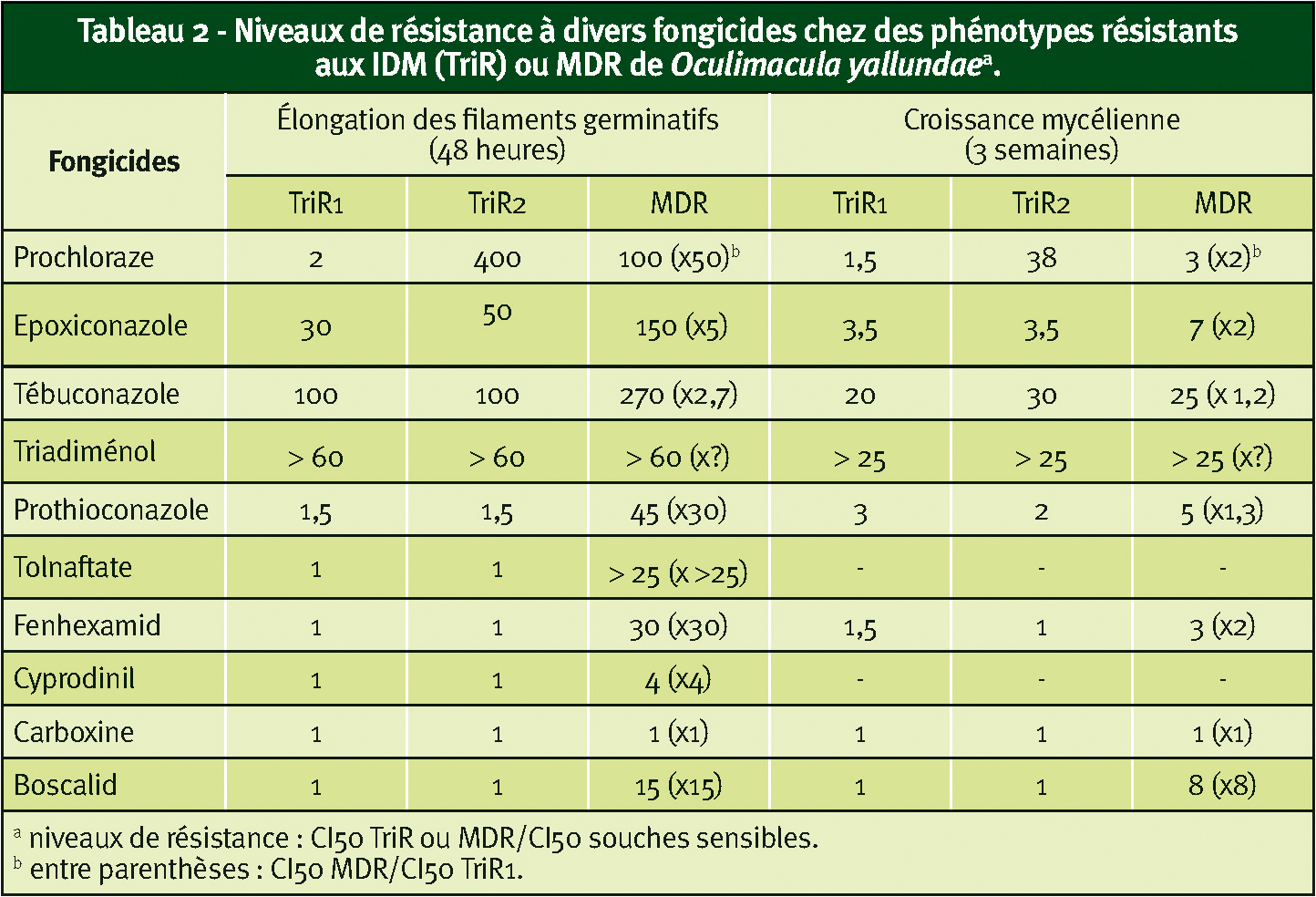
à partir du début des années 2000, la surveillance des populations françaises d'Oculimacula sp. nous a permis de détecter des souches présentant une résistance simultanée au boscalid et au prothioconazole.
Ce phénomène est plus prononcé dans les essais conduits sur spores que sur mycélium et il concerne aussi d'autres inhibiteurs stéroliques tels le tolnaftate et le fenhexamid, plus probablement le prochloraze et le cyprodinil.
Pour l'instant, cette résistance de type MDR n'a pas été observée chez O. acuformis (type lent ou « souches lentes »). Au vu de l'effet de certains triazoles (ex : triadiménol), elle s'est probablement développée chez O. yallundae (type rapide), espèce dominante en France, au sein du type TriR1 déjà résistant aux triazoles (Tableau 2).
En fait, ces souches MDR d'O. yallundae présentent un spectre de résistance proche des souches MDR2 de B. cinerea qui, comme précisé plus haut, impliquent la surexpression d'un transporteur à MFS.
En pratique, faibles fréquences
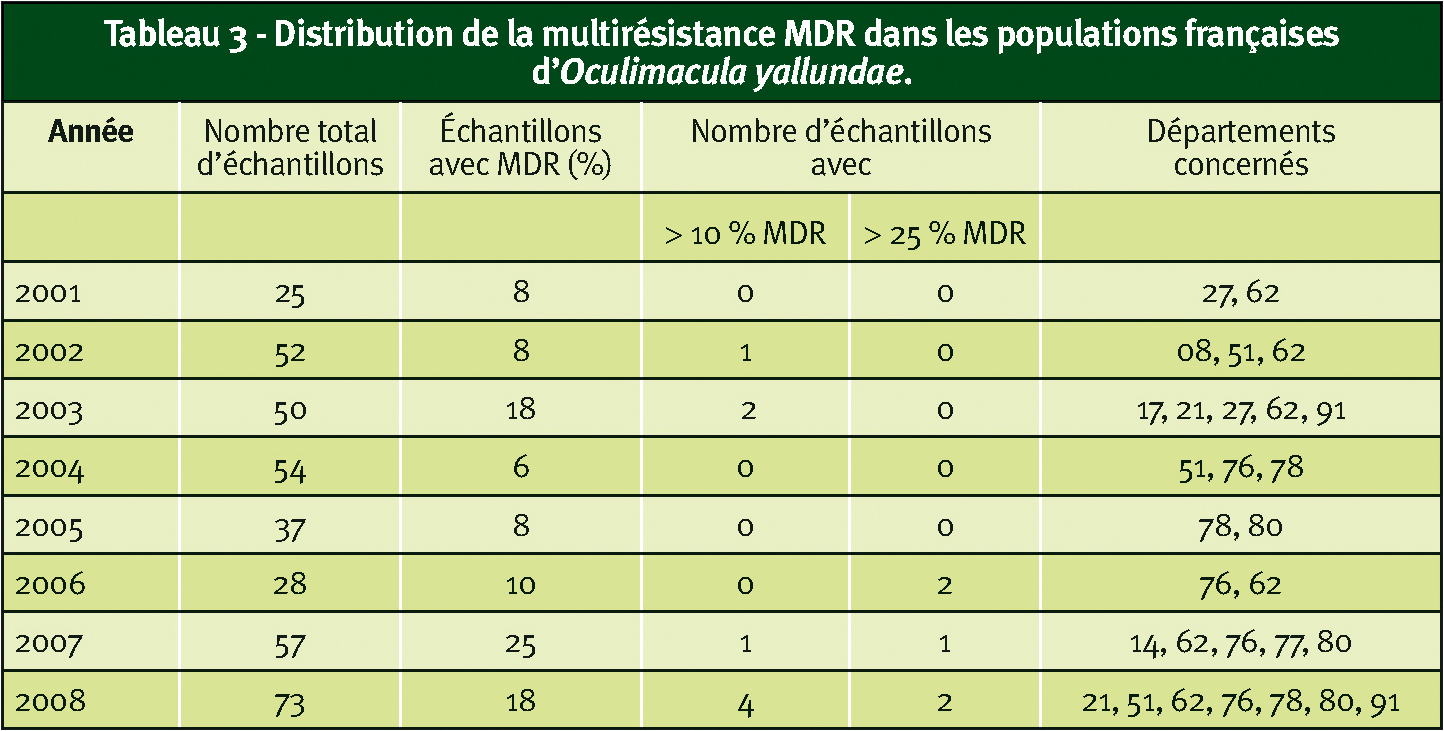
Ces souches MDR d'O. yallundae ont été détectées dans plusieurs régions françaises et, dans la plupart des cas, à des fréquences faibles. Les quelques échantillons présentant plus de 25 % de MDR venaient de sites d'essais pluriannuels traités au boscalid (nous ne disposons pas de telles modalités avec le prothioconazole).
À noter que, globalement, la situation est assez similaire pour la résistance spécifique aux anilinopyrimidines (AniR1) ; les quelques échantillons présentant plus de 25 % de souches AniR1 étaient issues de parcelles traitées au cyprodinil (Tableau 3).
Septoriose du blé
Six transporteurs en vedette
Pour la septoriose du blé, les investigations conduites en laboratoire montrent qu'au moins 5 ABC transporteurs (MgAtr1-MgAtr5) pourraient moduler la toxicité des IDM.
Toutefois, pour des isolats collectés au champ, aucune corrélation n'a été observée entre les réponses biologiques (niveaux de résistance), l'accumulation intracellulaire de fongicides et l'expression des ABC transporteurs.
Très récemment, l'implication d'un transporteur à MFS (MgMFS1) susceptible de moduler l'activité des IDM, des strobilurines et de molécules naturelles a également été démontrée en laboratoire. Les études réalisées sur des souches collectées au champ montrent que la plupart présentant une résistance spécifique aux strobilurines (avec changement G143A du cytochrome b) ont un niveau d'expression du MgMFS1 supérieur à celui des souches sensibles.
Il faut noter qu'aucune différence n'existe entre des souches fortement et très fortement résistances aux strobilurines et que, par ailleurs, certaines souches très fortement résistantes on un profil d'expression identique à des souches sensibles. Au vu de l'ensemble de ces résultats, il n'y a aucune démonstration formelle indiquant que chez M. graminicola la résistance aux IDM et/ou aux QoI serait polygénique chez des isolats collectés au champ.
Du nouveau depuis 2008
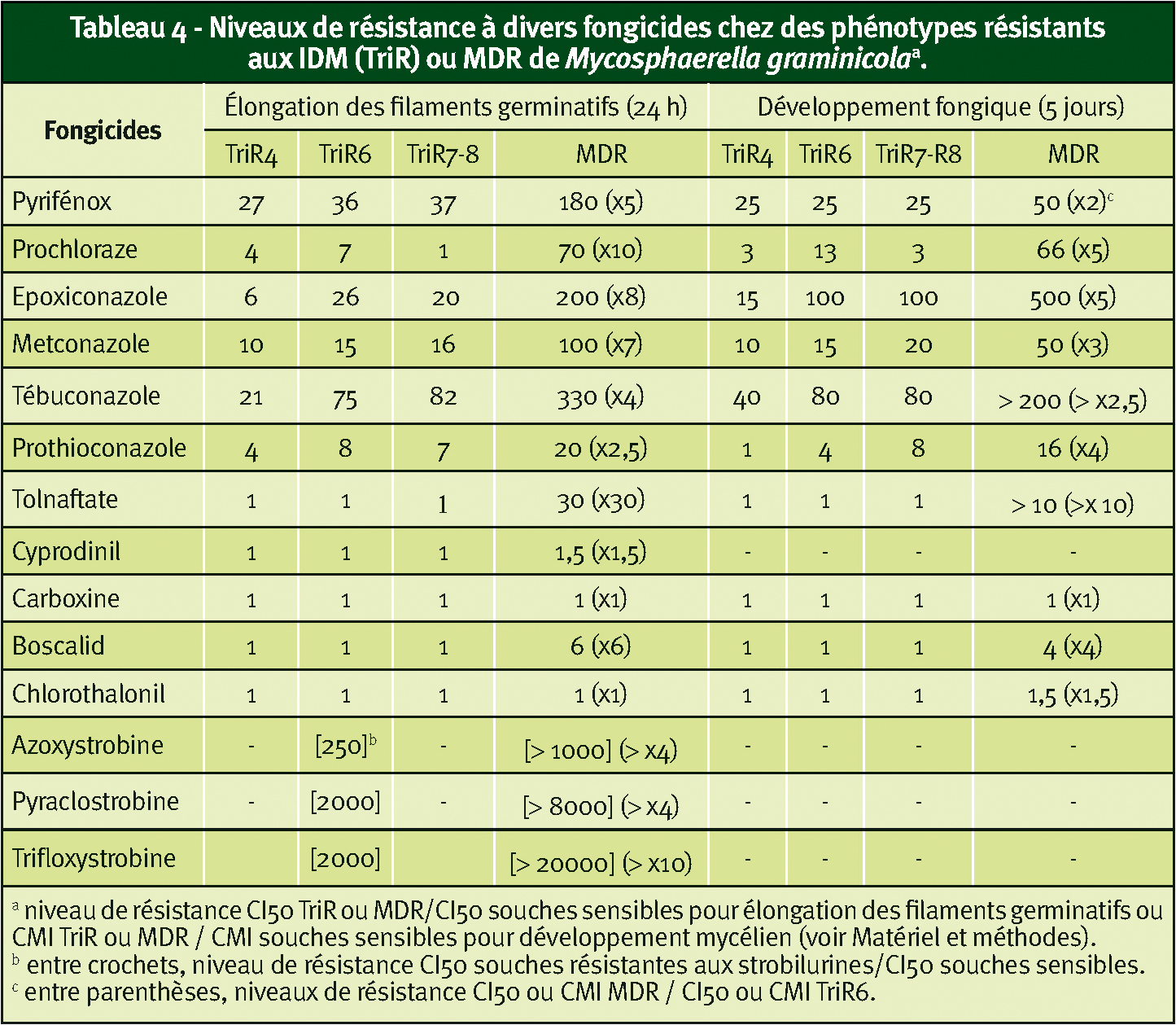
Or, en 2008 puis en 2009, nous avons détecté dans plusieurs régions céréalières françaises des souches de M. graminicola présentant un profil de résistance MDR.
À noter que tout d'abord elles possèdent des résistances spécifiques aux QoI et aux IDM déterminées par des mutations dans les cibles respectives de ces deux fongicides. Une de leurs premières caractéristiques, identique à celle observée chez B. cinerea et O. yallundae, est leur résistance au tolnaftate.
Il faut noter également pour ces trois parasites une sensibilité réduite vis-à-vis du boscalid mais pas de la carboxine (SDHIs). Concernant les strobilurines et les IDM, la résistance est augmentée d'un facteur de 2 à 10 par rapport aux souches présentant une résistance de cible de type TriR6 (Tableau 4). Des travaux sont en cours pour déterminer les bases moléculaires de cette MDR.
En pratique, là aussi faible fréquence
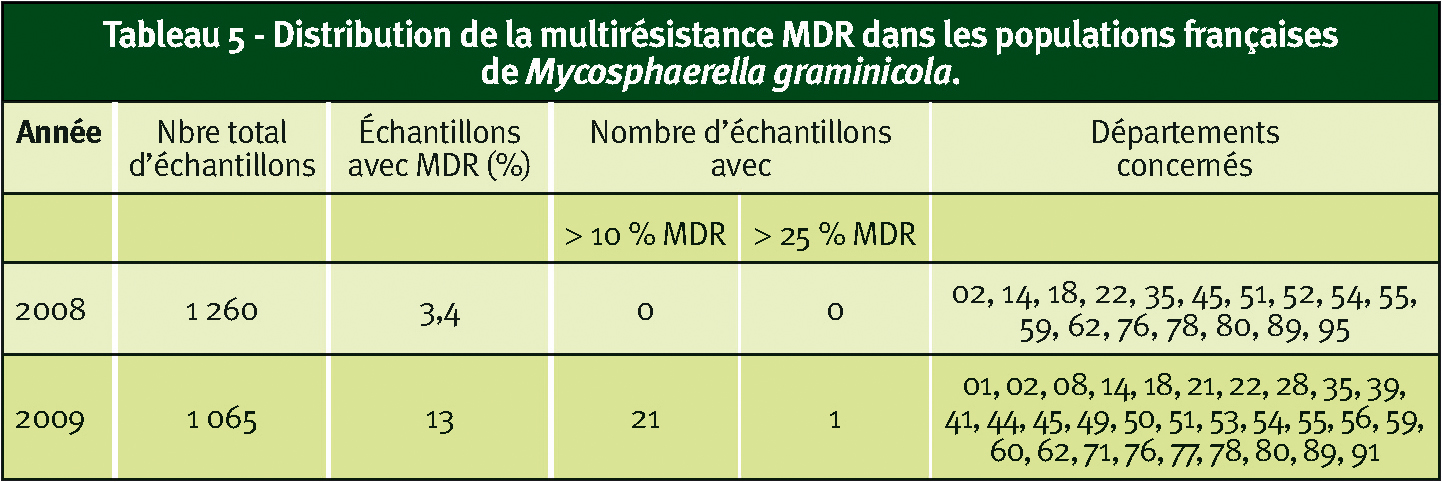
En 2008 et 2009, ces souches ont été décelées respectivement dans 3 % et 13 % des échantillons analysés dans notre laboratoire, indiquant une légère progression du phénomène. Toutefois, dans la majorité des cas, les fréquences de ces souches sont inférieures à 10 % et en 2009, sur un total de 1 065 échantillons analysables, 21 présentaient des fréquences supérieures ou égales à 10 % et seul un échantillon dépassait 25 %.
Ces souches MDR sont décelées surtout au nord de la Loire et en région Centre (Tableau 5).
Conclusion
MDR au champ, c'est prouvé
Grâce à une caractérisation phénotypique adaptée, basée principalement sur l'effet de divers fongicides sur l'élongation des filaments germinatifs émis par les spores, nous avons démontré l'existence de résistances MDR en conditions naturelles chez l'agent de la pourriture grise de la vigne, ainsi que ceux responsables du piétin-verse et de la septoriose du blé.
Chez B. cinerea, les bases moléculaires de cette MDR ont été caractérisées et impliquent la surexpression d'un transporteur de type ABC et/ou MFS ; ceux intervenant chez les pathogènes du blé ne sont pas connus.
En pratique, cela se complique
En pratique, au vu de la situation dans le vignoble champenois, il semble que ces résistances MDR seules aient un impact limité sur l'efficacité des fongicides anti-Botrytis.
En revanche, dans le cas de M. graminicola, il conviendra de déterminer pour les IDM (et peut-être un jour pour les SDHI) l'impact d'une sélection de souches combinant une résistance spécifique de cible et une MDR (situation de résistance polygénique).
Au laboratoire, pistes prometteuses
Enfin, des essais préliminaires réalisés au laboratoire sur B. cinerea et M. graminicola ont montré que certaines substances rétablissaient l'activité des fongicides. De tels modulateurs, dont certains sont développés dans le domaine médical, pourraient être associés à des fongicides agricoles. Ils se comportent comme les synergistes utilisés dans la lutte insecticide et qui sont des inhibiteurs d'enzymes de détoxification à P450.
Une autre approche pourrait consister à empêcher la fixation des fongicides sur les facteurs de transcription à l'origine de la surproduction des transporteurs. à noter que dans le domaine animal, cette approche pourrait également permettre de diminuer la pénétration intracellulaire et la détoxification des xénobiotiques car certains facteurs de transcription sont impliqués dans la régulation de gènes codant des transporteurs et des enzymes à P450.
<p>* INRA UMR BIOGER-CPP, Avenue Lucien-Bretignières 78850 Thiverval-Grignon walker@versailles.inra.fr</p> <p>(1) Pathogènes des plantes, sujets de la...</p> <p>(2) Conférence internationale sur les maladies des plantes organisée à Tours par l'AFPP, les 8 et 9 décembre 2009.</p>
Fongicides, résistances, mécanismes : cible, « détox » et transporteurs
La lutte chimique contre les champignons phytopathogènes est pratiquée depuis deux siècles. Mais elle s'est particulièrement développée après la seconde guerre mondiale avec l'avènement des fongicides organiques de synthèse. La première génération constituée de molécules multisites (ex : phtalimides, dithiocarbamates) a pris le relais de préparations minérales à base de cuivre et de soufre.
Face aux fongicides unisites...
À la fin des années 1960, de nouvelles matières actives performantes et systémiques sont apparues (ex : benzimidazoles, carboxamides). Cette nouvelle génération de fongicides agricoles, comme d'ailleurs tous ceux développés depuis lors (ex : phénylamides, IDM, QoI) sont des molécules unisites. Elles ont la propriété de se lier à une protéine impliquée dans un processus métabolique majeur (ex : respiration, division cellulaire, biosynthèse des stérols).
... des résistances par mutation de cible, parfois croisées
Avec un tel mode d'action, les champignons traités par un fongicide (ou une classe de fongicides) vont pouvoir s'adapter par l'intermédiaire d'une ou plusieurs mutations du gène codant pour sa protéine cible.
Précisons qu'il s'agit en général, non de la survenue de mutations nouvelles, mais de la sélection, par le (ou la famille de) fongicide(s), de souches portant la mutation et préexistantes aux traitements. Ce phénomène entrainant le changement d'un (ex : G143A sur le cytochrome b, la cible des QoI) ou plusieurs acides aminés (ex : chez CYP51, la cible des IDM) induit une réduction d'affinité du fongicide pour sa cible. Dans ces conditions, il y aura souvent résistance croisée entre toutes les molécules ayant le même mode d'action biochimique.
Parmi les autres mécanismes de résistance, il y a d'une part la détoxification accrue des substances et, d'autre part, une pénétration intracellulaire réduite.
Détoxification et pénétration intracellulaire réduite
Le premier mécanisme, peu fréquent chez cette classe de pesticides, se rencontre fréquemment, en revanche, pour les herbicides et surtout les insecticides. Parmi ces derniers, via notamment des mono-oxygénases à cytochrome P450, il peut y avoir résistance croisée entre des matières actives de mode d'action différents (ex : carpocapse du pommier).
Le second mécanisme, qui est corrélé à une moindre concentration intracellulaire des fongicides, résulte notamment de la production accrue de transporteurs membranaires capables d'excréter une grande variété de molécules toxiques. Cette résistance simultanée à plusieurs inhibiteurs est généralement dénommée MDR (Multidrug Resistance) et dans le cas de la levure, elle est qualifiée de PDR (Pleiotropic Drug Resistance).
Action des transporteurs, ce que dit la médecine
Ce phénomène a été décrit pour la première fois pour des antitumoraux chez les mammifères. Il est également observé dans le domaine médical, en conditions pratiques d'emploi, pour des antibactériens et antifongiques.
Deux types de transporteurs membranaires peuvent être impliqués : il s'agit de transporteurs à ABC (ATP-Binding Cassette) qui utilisent l'énergie libérée lors de l'hydrolyse de l'ATP, et des transporteurs MFS (Major Facilitator Superfamily) fonctionnant grâce au gradient électrochimique existant au niveau de la membrane cytoplasmique.
Chez la levure Saccharomyces cerevisiae et Candida albicans, un pathogène fongique humain, la production des transporteurs est sous la dépendance de facteurs de transcription qui se lient à la partie promotrice des gènes codant ces transporteurs. Chez les souches résistantes, la surproduction résulte soit de mutations dans le facteur de transcription, soit de modifications des promoteurs des gènes codant les transporteurs.
Par ailleurs, il a été montré récemment que les molécules fongitoxiques concernées par la MDR étaient capables de se fixer sur les facteurs de transcription.
Figure 1 - évolution de la résistance croisée (MDR, multidrug resistance) chez Botrytis cinerea dans le vignoble champenois.