Imprimer
Imprimer")
Botrytis cinerea (ph. Y. Elad)
 pour mettre en évidence les effets systémiques de l'application foliaire de sucres solubles sur la plante puis sur l'insecte. Photos 1 et 2 (vue générale et gros plan) : pulvérisation de la feuille 3 au stade 4-5 feuilles avec des solutions de sucres (D et L-glucose, D-fructose, saccharose à 0,1, 1 et 10 ppm) dés la préparation des solutions. Photo 3 : séchage des plantes après les traitements ; il dure environ une heure. Photos : S. Derridj")
Méthode utilisée sur maïs cultivé en serre (15 ± 1 °C la nuit et 25 ± 5 °C le jour) pour mettre en évidence les effets systémiques de l'application foliaire de sucres solubles sur la plante puis sur l'insecte. Photos 1 et 2 (vue générale et gros plan) : pulvérisation de la feuille 3 au stade 4-5 feuilles avec des solutions de sucres (D et L-glucose, D-fructose, saccharose à 0,1, 1 et 10 ppm) dés la préparation des solutions. Photo 3 : séchage des plantes après les traitements ; il dure environ une heure. Photos : S. Derridj
Parmi les sucres solubles, on trouve le « sucre » qui adoucit notre café, alias le saccharose, le fructose des fruits et le glucose très largement utilisé dans l'industrie agro-alimentaire. Ces sucres, pulvérisés à faibles doses sur les plantes, peuvent les aider à se défendre contre des bio-agresseurs : insectes, nématodes voire champignons pathogènes. Pas forcément en s'attaquant à ces agresseurs mais en perturbant leur reconnaissance de la plante comme « bonne à envahir ». Explications.
Le retrait du marché des substances phytopharmaceutiques (« pesticides ») les plus toxiques est en cours en Amérique du Nord et en Europe, en particulier en France. Les systèmes naturels de protection de la plante face aux agressions biotiques de son environnement figurent parmi les thématiques de recherche actuelle.
De nombreux laboratoires étudient la résistance des plantes aux bio-agresseurs et l'induction des systèmes de défense aux niveaux moléculaires, génétiques et physiologiques. Les défenses naturelles des plantes utilisent des voies de signalisation complexes souvent liées à celles de réponse aux stress abiotiques. Ceci expliquerait en partie que les inductions de défenses par des substances observées en laboratoire soient difficiles à reproduire au champ.
Un aspect moins étudié est celui de la résistance de la plante qui s'opère avant son attaque, lors de sa reconnaissance par le bio-agresseur. Nous le présentons ici.
Reconnaissance ou la voie des sucres
Utilisée par tous les vivants
Tous les organismes vivants utilisent les voies de signalisation par les sucres solubles.
La plante les met en œuvre pour intégrer les éléments internes et externes et maintenir à niveau constant certaines caractéristiques nutritionnelles, pour gérer les processus hormonaux de croissance et développement et pour articuler les réponses aux stress biotiques et/ou abiotiques (Koch, 2004 ; Rosa et al., 2009). Des centaines de gènes ont leur expression régulée par les sucres chez la plante.
Le comportement de l'insecte lui permet de sélectionner sa plante hôte en évitant les plantes défavorables. Les insectes lépidoptères comme la pyrale du maïs Ostrinia nubilalis et le carpocapse des pommes et des poires Cydia pomonella perçoivent par contact les sucres solubles à la surface des plantes comme des signaux influençant la reconnaissance de la plante et le dépôt de leurs œufs (Derridj et al., 1989 ; Lombarkia & Derridj, 2002 et 2008).
Les organismes moins mobiles (nématodes, micro-organismes phytopathogènes) ont aussi, malgré leur faible autonomie, des moyens de reconnaître la plante. Le mannose et le glucose exsudés par des sensilles sensorielles du nématode Meloidogyne incognita joueraient un rôle dans leur perception et orientation (Zuckerman & Jansson 1984 ; Perri, 1993). Le fructose agirait sur la germination du champignon Botrytis cinerea par des voies de signalisation (Doehlemann et al., 2005).
Trois équipes au travail
L'action de sucres solubles pulvérisés en infradoses (ppm) à la surface de plantes cultivées a été testée sur divers bio-agresseurs.
En France, l'INRA de Versailles l'a entrepris dans le but de modifier les signaux de surface de la plante (en particulier les sucres) utilisés lors de sa reconnaissance pour pondre par les lépidoptères.
Le Scottish Crop Research Institute (SCRI) (Grande-Bretagne) étudie les effets structuraux des sucres pour comprendre le mode d'induction systémique de la résistance de la tomate et de la pomme de terre aux nématodes par une molécule DMDP extraite d'une fève du Costa Rica. Analogue du fructose, le 2,5-dihydroxymethyl-3, 4-dihydroxypyrrolidine ou DMDP bloque la perception du nématode (Birch et al., 1993a).
Le centre de Volcani (Israël) étudie des méthodes de lutte biologique et culturale contre les micro-organismes phypathogènes par l'introduction de micro-organismes antagonistes dans le phylloplan et l'utilisation d'inducteurs de défense des plantes. L'étude des nutriments et des sucres déposés sur les plantes est exploitée pour augmenter l'activité et faciliter les introductions d'organismes utiles.
Nous allons évoquer les expérimentations menées en serre par ces trois équipes, qui démontrent l'induction systémique de résistances de la plante à des bio-agresseurs par des applications foliaires d'infra-doses de sucres solubles. Suivra le transfert de technologie en agriculture sur pommier contre C. pomonella.
Nous limiterons la description des effets à ceux du saccharose seul comparé à des témoins non traités. La compréhension des mécanismes par l'étude du métabolome (INRA Versailles et Université Paris Sud 11), des hormones végétales et des gènes régulés (SCRI) sont en cours.
Pulvérisation foliaire d'infra-doses de sucres, effet sur la pyrale du maïs
Essais sous abri à Versailles
La reconnaissance de la plante en tant qu'hôte et le dépôt d'œufs sur les feuilles par les femelles de la pyrale du maïs O. nubilalis sont influencées par la composition en métabolites primaires de la surface des feuilles, en particulier celle en sucres solubles. Ceux-ci, d'origine photosynthétique, traversent la cuticule et se trouvent en quantités de l'ordre du ng par cm² à la surface des feuilles (Derridj et al., 1996). Leur dynamique suit celle de la photosynthèse. Leurs proportions sont spécifiques et liées aux propriétés de perméabilité cuticulaire des espèces végétales (Stammitti et al., 1995).
Des infra-doses (entre 0,1 et 10 ppm) de sucres (D et L-glucose, D-fructose et saccharose, Produits Sigma-Aldrich) du même ordre quantitatif que celles existant naturellement à la surface des plantes (excepté le D-glucose, produit de synthèse) sont pulvérisées le matin sur la feuille 3 de maïs au stade 4-5 feuilles (photos 1 et 2). Les sucres et doses qui ont le plus d'effets sur la croissance de la plante ont été testés sur l'insecte : saccharose et L-glucose 10 ppm et D-fructose 0,1 ppm vs une plante non traitée avec le sucre.
Le travail est poursuivi en chambre climatisée (température 25 ± 1 °C, humidité relative 70-80 % et photopériode 16 h jour/8 h nuit). Les quatre cas sont testés sur la ponte d'O. nubilalis en conditions de choix en cages 6, 12 et 20 jours après la pulvérisation. Quatre plantes par cage sont présentées à une trentaine d'insectes de chaque sexe quelques minutes avant la scotophase. Le lendemain matin, on retire ces plantes des cages ; les pontes sont dénombrées sous loupe binoculaire et localisées sur la plante par niveau foliaire et par zone sur la feuille.
Résultats : action systémique du saccharose et du D-fructose
Seules les solutions de saccharose et de D-fructose respectivement à 10 ppm et 0,1 ppm ont un effet sur l'insecte et ceci après 20 jours. On observe des effets systémiques de réduction de la ponte de l'insecte de 30 et 43 % respectivement (n = 18, test de Bonferroni corrigé par Wilcoxon-Wilcox P < 0,05) sur l'ensemble de la plante, sans modification de sa répartition (sur variétés Anjou 258, LG 2447, Challenger, deux saisons de culture). Que le traitement soit effectué sur toute la plante ou seulement sur la feuille 3, les réductions de ponte sont similaires.
Un traitement unique de toute la plante par une solution de saccharose 10 ppm réduit après 20 jours d'environ 40 % la ponte d'O. nubilalis aux stades végétatifs du maïs doux variété Challenger. Deux traitements espacés de 20 jours à partir du stade 2 feuilles réduisent la ponte de 28 % (Figure 1a).
En revanche, un troisième traitement augmente les pontes de 80 % au stade grain laiteux du maïs (toujours Figure 1a). Mais ceci est dû en grande partie à une augmentation des pontes sur les faces supérieures des feuilles (+ 50 %). Or ce changement de comportement de l'insecte qui pond en général sur la face inférieure des feuilles défavorise l'éclosion des œufs qui sont plus exposés aux rayonnements solaires et aux intempéries (Pierre, 2007).
Essai au champ
En champ de maïs doux (semis tardif de la variété Challenger) l'ajout de solutions de saccharose 100 ppm à un traitement pyréthrinoïde tous les 14 jours minimum à partir du stade 2 feuilles confirme cette inversion d'effets sur les pontes entre les stades végétatifs et reproducteurs. Mais elle est moins marquée qu'avec le sucre seul en expérimentations en serre.
Dans les parcelles traitées au saccharose, les pontes, plus tardives et plus petites (nombre d'œufs par ooplaques), ont donc potentiellement moins de chenilles (figure 1b, page 11).
Sur un échantillonnage faible de 25 cannes de maïs par modalité à la récolte, on observe dans les parcelles additionnées de saccharose 100 ppm une réduction de 20 % des dégâts totaux à la récolte et une absence de dégâts sur l'épi principal vs 8 % sur le maïs traité avec l'insecticide seul.
Effet systémique contre les nématodes
Tomate sous abri en Écosse
Le D-glucose, le D-fructose et le saccharose appliqués en solution de l'ordre de la ppm le matin par pulvérisation foliaire ou par arrosage du sol induisent une résistance systémique de plantules de tomate au nématode Meloidogyne javanica (photos 4 et 5). On observe ainsi jusqu'à 60 % de réduction des galles sur racines. L'application foliaire est celle qui réduit le plus les dégâts et représente un avantage certain pour son application en cultures.
Pomme de terre sous abri en Écosse
Ces résultats sont du même ordre que ceux obtenus en Ecosse avec le DMDP en application foliaire sur tomate (Birch et al., 1993a) et contre les nématodes à kystes de la pomme de terre Globodera rostochiensis (Birch et al., 1993b). Dix semaines après la pulvérisation foliaire, le développement des galles et kystes et le nombre d'œufs sont réduits par le DMDP.
La variété de pomme de terre Heather génétiquement partiellement résistante présente 81 % de moins d'œufs de nématode par g de sol comparée à la variété sensible Désirée.
La dose de 100 µg/ml de DMDP réduit sur Heather de 80 % le nombre d'œufs du nématode par g de sol et de 96 % si on la compare à la variété Désirée. La combinaison de la résistance partielle avec le DMDP rend la variété Heather résistante et réduit les populations finales du nématode à 4 œufs par g de sol vs 106 sur la variété sensible non traitée (Birch et al., 1993 a).
Effets sur la contamination foliaire de la tomate par Botrytis cinerea
Sous serre en Israël
Deux à trois semaines avant le début des expériences avec les sucres, des plants de tomates de la variété 1402 (Hazera Genetics) sont cultivés en serre indemnes de maladies jusqu'au stade 5-nœuds.
Le saccharose à trois concentrations est pulvérisé sur les feuilles trois jours avant l'inoculation par le pathogène, tout en évitant de pulvériser le support et le substrat, et les plants sont comparés aux témoins non traités par les sucres. Au stade 5-nœuds, sept feuilles sont inoculées sur la plante avec 20 µL d'une suspension de conidies de B. cinerea (isolat BcI16), une goutte par feuille. Puis les plantes sont incubées à 20 ± 1 °C, 5150 lux, et 97 ± 3 % d'humidité relative.
Les conidies de B. cinerea sont obtenues à partir de cultures sur agar et dextrose de pomme de terre à 20 °C au bout de 10-14 jours.
La maladie est visible 3-5 jours après le dépôt de la suspension sur les feuilles. L'importance de chaque lésion est mesurée à l'aide d'une échelle visuelle basée sur la surface relative de la lésion par rapport à la surface maximum observée sur les témoins (= 113 mm²) et numérotée de 0, 1, 2, 5, 10, 20, 40, 75 à 100 %. La sévérité de la maladie est notée le premier jour après l'inoculation puis 7 et 10 jours plus tard et analysée avec le test Fisher PLSD P<0,05.
Résultats
Les doses de 1 et 100 ppm de saccharose déposées avant l'infection réduisent les lésions de 63 et 100 % respectivement par rapport aux lésions maximales observées sur les témoins (Figure 2).
Transferts de la technologie en agriculture
Applications foliaires en vergers de pommiers contre le carpocapse : Golden et Gala en Europe
Des tests de l'application du saccharose à 10 et 100 ppm ont été poursuivis par la société Anadiag sur deux variétés : Golden Delicious et Mondial Gala dans plusieurs vergers installés d'Europe (sud de la France, Italie, Grèce) pendant quatre années.
Quels que soient les niveaux d'infestation, le sucre seul a une efficacité Abbott ([(% de pommes attaquées sur le témoin – % pommes attaquées avec le traitement)/% de pommes attaquées sur le témoin]*100) sur les dégâts sur fruits à la récolte comprise entre 19,5 % et 63,3 % avec une moyenne de 40,6 ± 16 % (Ferré et al., 2008).
Golden et Anna en Algérie
Des essais similaires ont été effectués en Algérie (2009) sur les variétés Golden Delicious et Anna. La solution de saccharose (Sigma, référence S1888 sup. 99,5 %) est utilisée seule à une concentration de 100 ppm (10 g pour 100 litres) dans deux vergers installés de 20 ha situés à Ain Djasser (Batna) et Yabous (Khenchela). Les applications foliaires sont pratiquées avec un pulvérisateur à dos. Le volume de liquide est adapté au développement végétatif du verger afin d'assurer un bon mouillage de la végétation conformément aux bonnes pratiques agricoles (compris entre 600 et 1 000 l/ha selon la taille des arbres). Les essais reposent sur un plan expérimental en blocs de Fisher randomisés avec 4 répétitions. Toutes les modalités sont réparties de manière aléatoire à l'intérieur de chacun des blocs. Les traitements commencent en avril dès le début des captures du carpocapse par piège à phéromones. Ils sont pratiqués tous les 20 jours tôt le matin jusqu'à la fin de la 3e période de ponte.
Les taux d'infestation par le carpocapse sur les arbres témoins sont élevés sur les deux variétés (environ 40 %). Les efficacités Abbott du saccharose 100 ppm sur les dégâts du carpocapse à la récolte des pommes sont de 59,8 ± 11,6 sur la variété Golden Delicious et de 48,9 ± 6 sur la variété Anna (Figure 3). Les résultats confirment ceux enregistrés auparavant par la société Anadiag dans les pays européens.
Risques d'effets sur d'autres ravageurs non ciblés
Acariens du tilleul
Un changement de composition des substances nutritionnelles ou de métabolites secondaires dû à la pollution des feuilles du tilleul favorise les acariens (Heinrich, 1988).
Le rapport entre les sucres solubles totaux et les composés phénoliques augmente dans les feuilles polluées vs les témoins en raison d'une diminution des concentrations des composés phénoliques. Tomczyk et al., (1997) indiquent que la concentration en sucres simples (fructose, glucose) est plus importante (9,4 g/g contre 7,2) dans les feuilles de Tilia platyphyllos les plus attaquées par Eotetranychus tiliarium. L'impact d'une modification du phylloplan en métabolites due à l'apport exogène de sucres mérite d'être vérifié sur les acariens phytophages et auxiliaires.
Une année de test sur pommiers
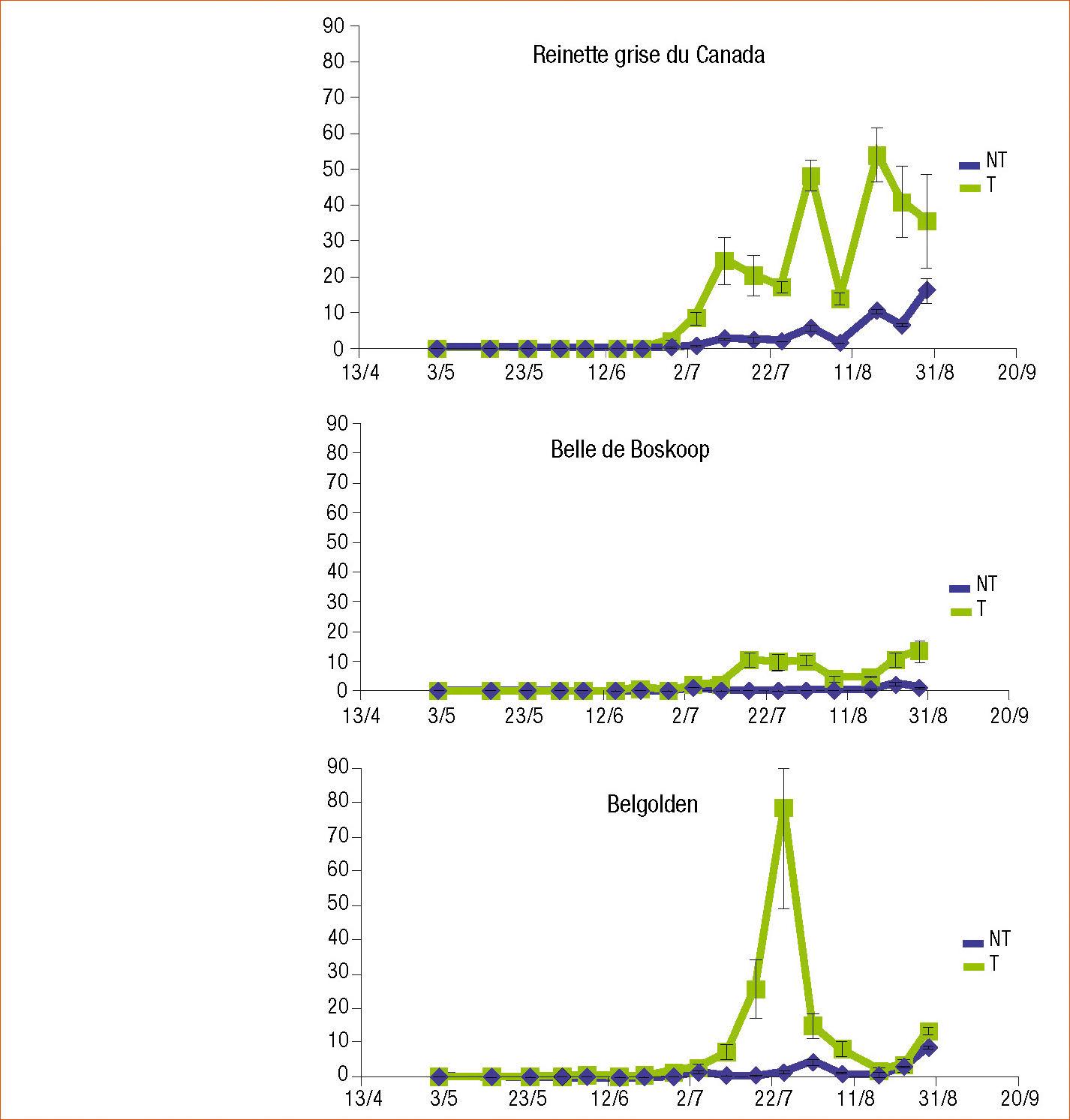
Les effets du traitement foliaire avec une solution de saccharose 10 ppm à une fréquence de 20 jours ont été examinés sur l'acarien tétranyque : Panonychus ulmi et ses acariens prédateurs, les phytoséiides, sur 3 variétés de pommier : Reinette grise du Canada, Belle de Boskoop et Belgolden. Les acariens phytophages et auxiliaires sont dénombrés sur 20 feuilles/arbre. Le suivi des populations a été réalisé durant 4 mois.
Pour chacune des dates de relevés (12 arbres par variété, 3 blocs par variété), les variabilités des dénombrements ne permettent pas de conclure à des différences statistiquement significatives pour les variétés Reinette grise du Canada et Belle de Boskoop (Wilcoxon). En revanche le traitement par le sucre a significativement favorisé l'acarien phytophage sur la variété Bel Golden (Wilcoxon P < 0,05). Il semble que globalement sur la saison certaines variétés soient plus sensibles aux acariens et que l'application de saccharose 10 ppm a augmenté les populations d'acariens phytophages proportionnellement aux sensibilités naturelles observées sur les témoins (Figure 4).
Les suivis des populations d'acariens auxiliaires n'ont montré en revanche aucune différence significative entre pommiers traités au saccharose 10 ppm et ceux non traités, pour chacune des trois variétés étudiées. Les augmentations de populations des phytophages n'ont pas eu d'incidence sur les populations d'auxiliaires.
Autre année, autre tendance
Les essais ont été reconduits l'année suivante sur les mêmes arbres selon le même protocole. Aucun impact du traitement saccharose 10 ppm n'est cette fois-ci mis en évidence sur les populations d'acariens phytophages et d'auxiliaires.
Nous avons donc constaté sur la variété Belgolden un effet négatif de l'apport exogène de saccharose 10 ppm favorisant l'acarien phytophage. Mais cet effet non ciblé semble plutôt irrégulier par rapport aux effets du saccharose 10 ppm sur le carpocapse très réguliers enregistrés sur le pommier plusieurs années.
On ne peut toutefois l'exclure et cela montre la nécessité d'études larges des agro-écosystèmes en ce qui concerne l'examen des effets des sucres sur la plante et ses bio-agresseurs. Il s'agira d'apprécier les équilibres obtenus en pesant les enjeux économiques et écologiques.
Conclusion
Nous montrons par l'expérimentation que la pulvérisation foliaire d'infra-doses de l'ordre de la ppm de sucres simples induit une réaction de la plante qui se traduit par une résistance de celle-ci face à plusieurs bio-agresseurs étudiés. Les mécanismes impliqués, complexes, se manifestent à la fois avant l'attaque, au moment de la reconnaissance de l'hôte par l'agresseur à la surface de la plante, et lors de l'attaque par des voies de défenses activées dans la plante (publication et recherche en cours).
L'efficacité sur les dégâts pourrait donc être expliquée d'une part par un abaissement des populations qui attaquent la plante et d'autre part par une mortalité accrue lors de l'attaque. Les effets de résistances sont spécifiques du couple variété de plante-bioagresseur. En effet, selon l'ENPS, certaines variétés de pommiers comme Jonagold ne montrent pas d'amélioration du traitement chimique contre le carpocapse par l'addition de saccharose.
L'utilisation des sucres peut permettre d'abaisser les taux de populations des bio-agresseurs à des seuils plus faciles à combattre ou au-dessous des seuils économiques, et ainsi participer à augmenter l'efficacité des méthodes de protection intégrée ou d'agriculture biologique. La sauvegarde de la diversité biologique engendrée par ces différents facteurs reste toutefois à surveiller et étudier dans des agro-écosystèmes. On ne peut totalement exclure des effets favorisant d'autres bio-agresseurs ou défavorisant des auxiliaires.
La résistance induite peut être aussi efficace (efficacité Abbott) sur la réduction des dégâts que la pulvérisation d'un insecticide chimique (Ferré et al., 2008). L'addition d'infra-dose de sucres à un pesticide chimique ou biologique peut permettre de réduire sa dose et/ou d'augmenter son efficacité.
Le mode d'action de la pulvérisation foliaire des sucres solubles en infra-doses est complexe ; afin d'optimiser ses effets, il est nécessaire d'approfondir les recherches pour en comprendre les mécanismes. Une des hypothèses est celle du déclenchement d'une action rapide par pénétration dans les cellules de garde des stomates. La voie de signalisation du fructose, qui semble être celle qui a les effets les plus généraux d'induction en serre, commence à peine à être étudiée chez les plantes et dans les cellules de garde en particulier. Les systèmes mis en jeu s'échelonnent dans le temps et s'expriment en cascade en faisant intervenir de plus en plus de voies de signalisation et de gènes.
Une ouverture prometteuse de recherche s'offre devant nous. Son exploitation dans la sélection variétale ou la biotechnologie serait aussi très économique en termes d'efforts de recherche.
Les sélectionneurs et les producteurs de fruits et légumes pourraient, grâce à la connaissance des mécanismes et des gènes concernés, bénéficier de la création de variétés dont la résistance constitutive ou partielle serait accrue. Une variété actuelle partiellement résistante peut le devenir à 100 % après une application de sucre sur la plante ou en arrosage du sol (pomme de terre/nématode). Induire une résistance partielle est maintenant considérée comme commercialement utile dans la lutte intégrée, qui constitue une exigence de l'UE d'ici 2014.
<p>* INRA (Institut national de recherche agronomique), UMR 1272, Physiologie de l'insecte, 78000 Versailles.</p> <p>** CETU Innophyt, Université François-Rabelais, de Tours, avenue Monge, 37200 Tours.</p> <p>*** SCRI, Dundee, DD2 5DA Écosse, Royaume Uni.</p> <p>** ** ARO, Plant Pathology and Weed Research, Volcani Center, Bet Dagan 50250, Israël.</p> <p>*** ** Université de Batna, Faculté des Sciences,</p> <p>Département d'Agronomie, Laboratoire d'amélioration de la production agricole et de la protection des écosystèmes en zones arides. 05000 Batna, Algérie.</p>
Figure 1 - Action sur les pontes d'Ostrinia nubilalis de la pulvérisation d'une solution de saccharose à 10 ppm sur maïs doux (var. Challenger) d'une solution de saccharose 10 ppm sous serre (a) et en champ (b) :
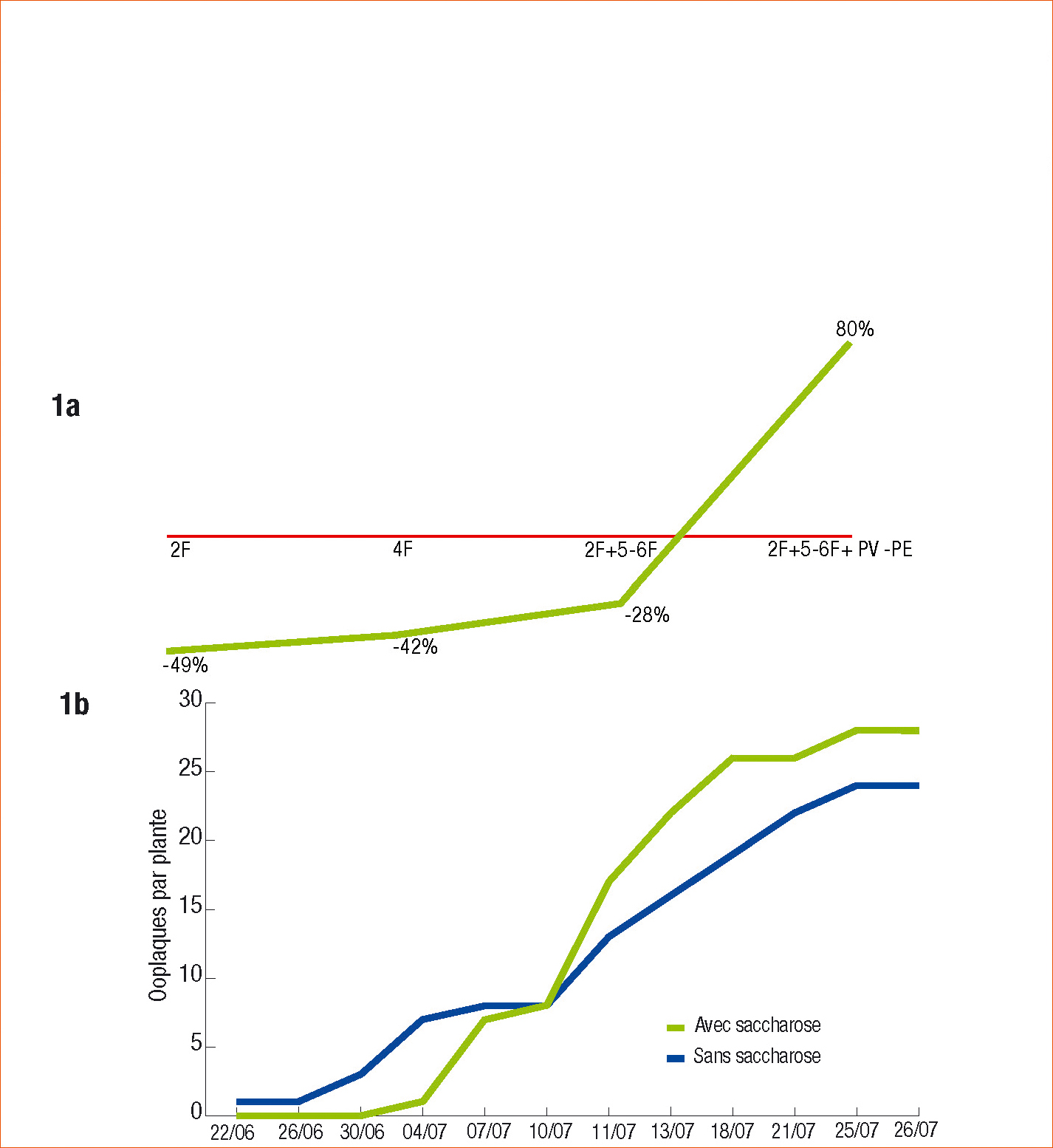
1a : 2 fois 8 plantes données en condition de choix à l'insecte vs des plantes témoins pulvérisées avec de l'eau. Les pontes sont estimées sur les plantes 20 jours après un unique traitement aux stades 2 et 4 F, et 20 jours après le dernier de 2 ou 3 traitements à partir du stade 2 F (test de Wilcoxon p < 0,10) (Pierre, 2007).
1b : Dénombrements des pontes sur maïs cultivé dans une parcelle de 4 ha (Spuis France 45) réalisés 2 fois par semaine au cours de la saison, représentés en variables cumulées. Traitements avec un pyréthrinoïde seul ou additionné de saccharose 100 ppm (bandes de maïs de 30 rangs sur 24 m alternées et répétées 3 fois, 100 plantes par répétition prises au hasard). Traitements commencés au stade 2 feuilles (2F) tous les 14 jours au minimum sur toute la saison (Programme Innophyt, 2007, FREDON Centre/Terr' Loire).
")
Photo 4 - Galle de Meloidogyne spp sur racines de tomate. Photos : A. Nicholas E. Birch (SCRI)

Photo 5 - Effets du saccharose 1 ppm sur les racines de tomates contaminées par les nématodes à galles Meloidogyne spp en comparaison avec un témoin eau.
Figure 2 - Effets, sur la sévérité de l'infection de la tomate par Botrytis cinerea, de l'application foliaire de saccharose aux concentrations de 0, 0,1, 10, 100 µg/ml.
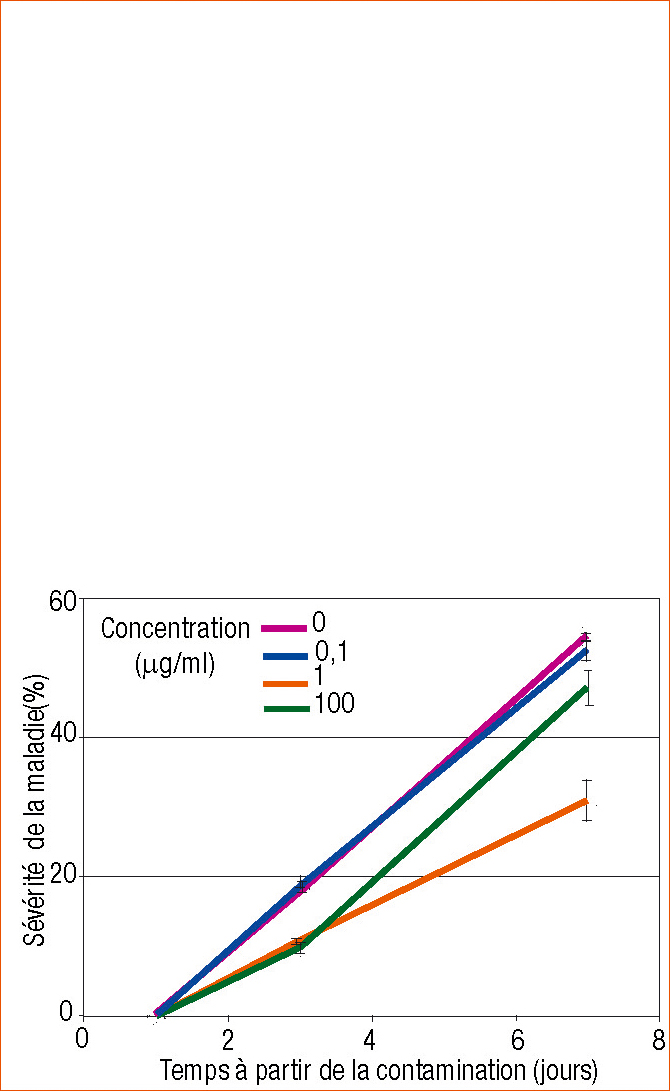
Trois jours avant la contamination, les plantes sont pulvérisées avec une suspension de 5 × 105 cellules pathogènes/ml, supplémentées par 0,1 % de D-glucose et 0,1 % de KH2PO4. La sévérité des lésions est exprimée en pourcentage par rapport au maximum de surface de lésion (= 113 mm²) sur les témoins. Chaque moyenne de 5 répétitions (1 répétition = 1 plante) est représentée avec une barre d'erreur standard.
Figure 3 - Taux d'infestations des pommes par le carpocapse et efficacités Abbott du traitement saccharose 100 ppm sur les dégâts sur fruits dus aux chenilles de carpocapse à la récolte. Deux variétés de pommier Golden Delicious et Anna en vergers installés, 4 blocs de 3 arbres pour Golden Delicious et de 2 à 1 arbre pour Anna, 125 fruits examinés sur Golden Delicious et 200 à 150 sur Anna (test de Wilcoxon P < 0,05).
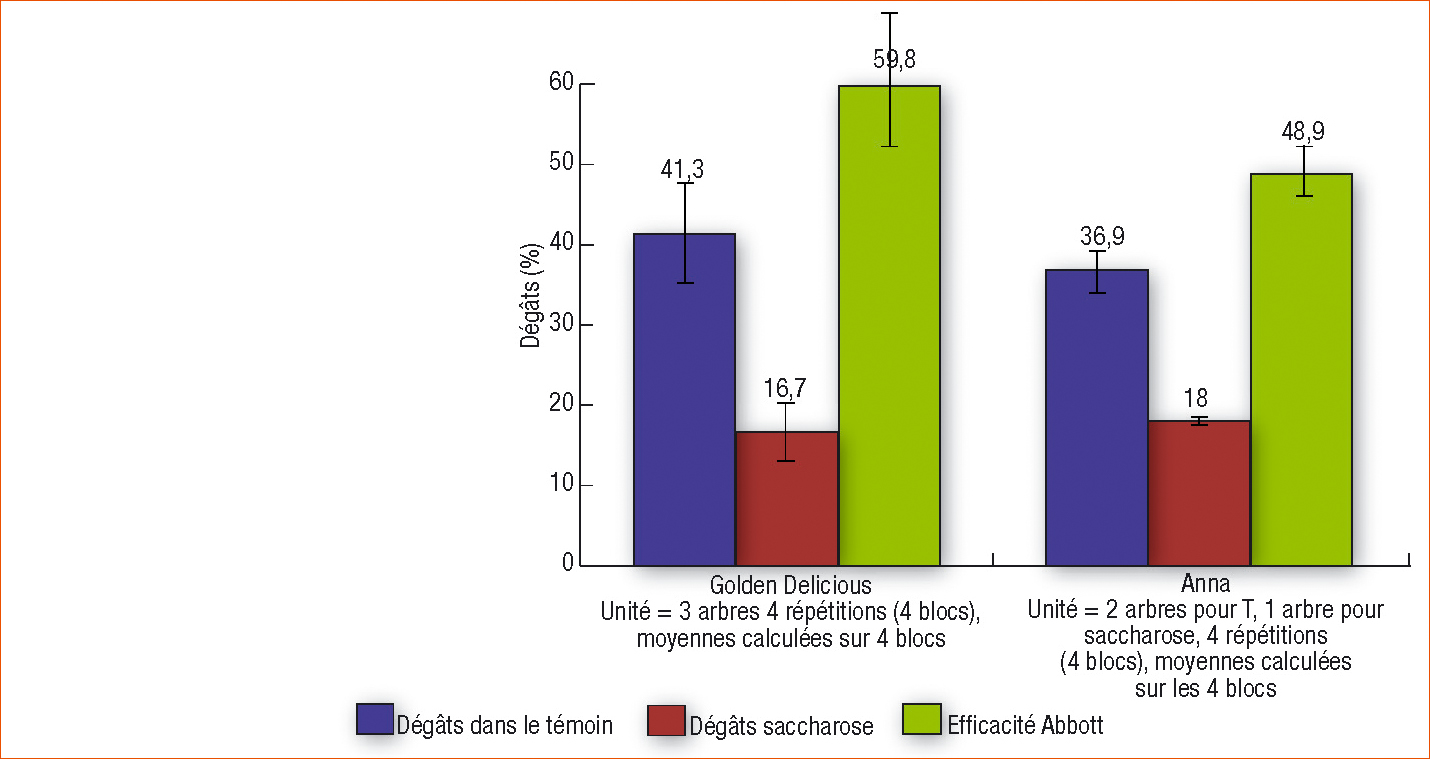
Figure 4 - Populations d'acariens ravageurs au cours de la saison 2006 sur pommiers d'ornement cultivés au jardin du château de Villandry selon les modalités témoin non traité au saccharose (NT) et traitée par le saccharose 10 ppm (Sac) sur trois variétés de pommier (Innophyt).