 Imprimer
Imprimer
La protection du blé utilisait des fongicides inhibiteurs du complexe III. Ce mode d'action a été compromis par la sélection de souches très résistantes. Photo : M. Decoin
 est une cible des inhibiteurs du complexe III que sont l'acéquinocyl et le bifénazate... Mais des résistances ont été sélectionnées. Photo : A. Migeon - Inra")
L'acarien ravageur Tetranychus urticae (ici, une femelle) est une cible des inhibiteurs du complexe III que sont l'acéquinocyl et le bifénazate... Mais des résistances ont été sélectionnées. Photo : A. Migeon - Inra
, un QoI-D (amétoctradine) et un QiI (cyazofamide). Photo : Pixabay")
Sur la vigne, il existe plusieurs fongicides inhibiteurs du complexe III : des QoI-P (azoxystrobine, pyraclostrobine, krésoxim-méthyl), un QoI-D (amétoctradine) et un QiI (cyazofamide). Photo : Pixabay
 ou acaricide (acéquinocyl). Photo : Decoin")
Sur pommier, des inhibiteurs du complexe III sont autorisés comme fongicides (krésoxim-méthyl, pyraclostrobine) ou acaricide (acéquinocyl). Photo : Decoin

Tableau 1 : Classification et propriétés des inhibiteurs du complexe mitochondrial III

Tableau 2 : Résistances aux inhibiteurs de type QoI liées au cytochrome b
Les substances « inhibiteurs respiratoires du complexe III » sont utilisées en phytopharmacie mais aussi en pharmacie vétérinaire et humaine, notamment contre le paludisme. Elles sont en butte à des phénomènes de résistance de certains des bioagresseurs qu'elles visent. Parmi elles, plusieurs fongicides agricoles sont concernés.
Pour savoir gérer ces phénomènes de résistance, il faut d'abord comprendre leurs mécanismes et en faire l'inventaire. C'est ce que propose le présent article.
Situer les inhibiteurs du complexe III
Phase terminale de la respiration cellulaire
Chez les eucaryotes (organismes dont les cellules ont un noyau et des mitochondries), la respiration cellulaire permet la production d'énergie sous forme d'ATP (adénosine tri-phosphate) et résulte du catabolisme oxydatif de sucres, lipides ou protéines.
Les phases terminales s'opèrent au niveau des mitochondries via le cycle de Krebs et la chaîne respiratoire. Celle-ci comporte quatre unités fonctionnelles (ou complexes mitochondriaux), intégrées dans la membrane mitochondriale interne :
- la NADH-ubiquinone réductase (complexe I) ;
- la succinate-ubiquinone réductase ou succinate déshydrogénase (complexe II ou SDH) ;
- l'ubiquinone-cytochrome c réductase (complexe III) ;
- la cytochrome c oxydase (complexe IV) assurant la réduction de l'oxygène en eau.
Le transfert des électrons entre ces complexes implique le coenzyme Q et le cytochrome c.
Le gradient de protons engendré par le fonctionnement de la chaîne respiratoire permet, via une ATP-synthase (complexe V), la synthèse de l'ATP, qui est libérée dans le cytoplasme et contribue aux différentes fonctions cellulaires (Figure 1 page suivante).
Substances agricoles
Plusieurs familles de fongicides et acaricides agricoles inhibent la respiration, notamment en stoppant la fixation du coenzyme Q sur les complexes I, II ou III.
Les inhibiteurs du complexe I sont principalement des acaricides de type Meti (Figure 1) dont le fénazaquin, le fenpyroxymate, le pyrimidifen et le tolfenpyrad. Ce dernier, ainsi que le diflumétorim (analogue structural du pyrimidifen) sont utilisés comme fongicides au Japon.
Les inhibiteurs du complexe II comptent une famille majeure de fongicides, les SDHI (Figure 1) ou carboxamides (bixafen, boscalid, carboxine, fluopyram, fluxapyroxad, isopyrazam, penthiopyrad...). S'y ajoutent deux proacaricides récents de type β-cétonitrile, le cyénopyrafen et le cyflumétofen.
Pour leur part, les inhibiteurs du complexe III seront détaillés plus loin.
Tous ces inhibiteurs respiratoires ont en commun d'être des unisites, susceptibles d'être en butte à des phénomènes de résistance, suite à des modifications de leurs cibles respectives ou encore via d'autres mécanismes (Leroux & Walker 2010a, b).
L'objet de cet article est d'apporter les dernières informations disponibles concernant les inhibiteurs du complexe III et en particulier l'amétoctradine, et de préciser leurs risques de résistance respectifs et les stratégies de gestion adaptées.
Structure et fonctionnement du complexe III
Rôles des cytochromes b et c1, de l'ISP...
Au niveau de la chaîne respiratoire mitochondriale, le complexe III ou cytochrome bc1 catalyse le transfert des électrons du coenzyme Q (dont les formes oxydées et réduites sont respectivement l'ubiquinone UQ et l'ubiquinol UQH2) au cytochrome c.
Chez les eucaryotes, ce complexe III comporte une dizaine de peptides. Parmi eux, le cytochrome b (seule sous-unité codée par un gène mitochondrial), la protéine fer-soufre Rieske (ISP) et le cytochrome c1 jouent un rôle essentiel. Ainsi, les électrons sont transférés via des atomes de fer inclus dans deux hèmes du cytochrome b (bl et bh), dans un cluster [2Fe-2S] de l'ISP et dans un hème du cytochrome c1 (Figure 2).
L'apoprotéine du cytochrome b est insérée dans la membrane mitochondriale interne via huit hélices transmembranaires (HTM) notées A à H. Celles-ci sont reliées par des boucles localisées soit vers l'intérieur (boucles bc, de, fg), soit vers l'extérieur (boucles ab, cd, ef, gh) de la mitochondrie.
Fixation du coenzyme Q
Il existe deux sites de fixation du coenzyme Q au niveau de cytochrome b : l'un proche de la matrice mitochondriale et dénommé Qi (« i » pour intérieur), et l'autre orienté vers l'extérieur et dénommé Qo (« o » pour out, extérieur en anglais).
Le site Qi, où s'opère la réduction de l'ubiquinone, implique des acides aminés de la partie N-terminale du cytochrome b, dont l'hélice A (résidus 15 à 40), des parties C et N terminales respectivement des hélices D et E ainsi que de la boucle de (résidus 195 à 230).
L'oxydation de l'ubiquinol s'opère au site Qo. Les acides aminés concernés se situent d'une part dans la partie C terminale de l'hélice C et la boucle cd (résidus 125 à 155), et d'autre part dans la boucle ef et la partie N-terminale de l'hélice F (résidus 255-295) (Figure 2 ; Fisher and Meunier 2008).
Études récentes
Diverses approches génétiques, enzymologiques et biophysiques ont permis de mieux comprendre le fonctionnement du complexe III. Les sites de fixation du coenzyme Q ont été caractérisés.
La plupart de ces études ont été réalisées avec des complexes III de vertébrés. Mais BASF a récemment étudié un Pythium avec l'amétoctradine (Esser et al., 2004 ; Berry and Huang 2011 ; Fehr et al., 2014).
Qui sont les inhibiteurs du complexe III ?
À propos de certains antibiotiques
Plusieurs antibiotiques peuvent interagir avec le complexe III. L'élucidation de leur mode d'action a permis de mieux comprendre le fonctionnement du cytochrome b.
Ainsi, il a été établi que l'antimycine A et d'autres dilactones cycliques (produites par Streptomyces spp.) et l'ilicicoline H (produite par Cylindrocladium ilicicola) se fixent au niveau du site Qi, alors que le myxothiazole, la strobilurine A et la stigmatelline (produits respectivement par Myxococcus fulvus, Strobilurus tenacellus et Stigmatella aurantiaca) interagissent avec le site Qo.
Une analyse détaillée de cette seconde classe d'inhibiteurs a dévoilé que le myxothiazole et la strobilurine A sont localisés près de l'hème bl (autour de 6 Å), alors que la stigmatelline en est plus éloignée (autour de 10 Å) mais interagit avec la sous-unité ISP.
Classes : QiI, QoI-P, QoI-D et QioI
De ce fait, la communauté scientifique distingue trois classes principales d'inhibiteurs du complexe III : QiI, QoI-P (proche de l'hème bl) et QoI-D (distant de l'hème bl). Dans la nomenclature du Frac, aux inhibiteurs QoI-P et QoI-D correspondent respectivement les QoI/strobilurines (C3) et les QoSI (Quinone outside Stigmatellin binding type inhibitor ; C8).
La funiculosine et l'ascochlorine (produits par Ascochyta viciae et Penicillium funiculosum), pouvant se fixer au niveau des sites Qi et Qo, ajoutent une quatrième classe, les QioI (Esser et al., 2004)(Tableau 1).
Les QiI, fongicides agricoles, entre autres
Parmi les fongicides de synthèse interférant avec le complexe III, seules les diméthyl-sulfonamides anti-oomycètes (cyazofamide, amisulbrom) sont de type QiI. Leur action résulterait d'une fixation covalente de la sulfonamide dans la zone où l'ubiquinone est réduite. Au laboratoire, le diuron se comporte comme un inhibiteur de type QiI chez la levure Saccharomyces cerevisiae, mais son activité herbicide résulte de l'inhibition de la photosynthèse.
Plusieurs aza-naphthalènes dont le HQNO ou le HDQ (qui présente une activité antipaludique) sont des QiI. Certains, comme le NQNO, pourraient se fixer sur les sites Qo et Qi. De façon générale, les QioI se fixent dans la même zone que la stigmatelline au niveau du site Qo (Tableau 1).
Les QoI-D : médicaments contre le paludisme et la toxoplasmose, acaricides, fongicides
Plusieurs hydroxynaphthoquinones, dont l'atovaquone et l'acéquinocyl, présentent un comportement voisin de la stigmatelline (QoI-D).
L'atovaquone est utilisée contre le paludisme provoqué par Plasmodium sp. mais aussi la toxoplasmose et une pneumocystose provoquée par le champignon Pneumocystis jiroveci (Fisher and Meunier 2008).
Enfin, l'acéquinocyl est un proacaricide efficace notamment sur Tetranychus urticae (Van Nieuwenhuyse et al., 2009).
Toujours parmi les QoI-D, il est possible de mentionner d'une part des dioxobenzothiazoles expérimentales (HHDBT, UHDBT), d'autre part l'amétoctradine, détaillée dans l'encadré ci-dessus.
Cette triazolopyrimidine (dont la structure chimique s'apparente à celle d'antimicrotubules utilisés comme anticancéreux) a été récemment introduite contre divers oomycètes dont les mildious de la vigne et de la pomme de terre (Fehr et al., 2014).
Il faut noter la présence d'une longue chaîne aliphatique hydrophobe chez la plupart de ces QoI-D synthétiques.
Enfin, l'acaricide bifénazate a été initialement décrit comme neurotoxique mais l'existence d'une résistance croisée avec l'acéquinocyl suggère qu'il s'agit plutôt d'un QoI-D, probablement sous la dépendance de son dérivé diazénique (Van Nieuwenhuyse et al., 2009).
Les QoI-P, fongicides strobilurines, d'autres fongicides et quelques acaricides
Les QoI-P synthétiques les plus nombreux sont des strobilurines (Tableau 1). Certaines comme l'azoxystrobine, la picoxystrobine (fongicides) ou la fluacrypyrime (acaricide) renferment le même toxophore que la strobilurine A : le méthoxyacrylate. D'autres fongicides polyvalents sont des méthoxy-iminoacétates (krésoxim-méthyl, trifloxystrobine), des méthoxy-iminoacétamides (dimoxystrobine), des méthoxy-carbamates (pyraclostrobine) ou des dioxazines (fluoxastrobine).
La famoxadone (oxazolidinedione) et la fénamidone (imidazolinone) sont deux QoI très efficaces contre les oomycètes. Leur site de fixation recouvre partiellement celui de la stigmatelline A et des strobilurines. La proximité de l'hème bl et la résistance croisée avec les strobilurines pour la substitution G143A dans le cytochrome b les fait classer comme des QoI-P.
Enfin, le pyribencarb (benzylcarbamate) présente une similitude structurale avec les strobilurines mais la faible résistance chez les mutants G143A et la toxicité réduite sur les complexes III animaux et végétaux suggèrent un mode de fixation particulier au niveau du site Qo (Fernandez-Ortuno et al., 2008 ; Leroux and Walker 2010b ; Kataoka et al., 2010 ; Berry and Huang 2011)(Tableau 1).
Chevauchements QoI-P/QoI-D
Les analyses cristallographiques conduites sur des complexes III mitochondriaux de vertébrés indiquent que les domaines de fixation des QoI-D et QoI-P se chevauchent.
Ainsi, pour le cytochrome b, les résidus G143, I147, P271, E272 et F275 sont proches de tous les QoI. Parmi ces acides aminés, le glutamate E272, inclus dans le motif PEWY et essentiel à l'activité catalytique, interagit avec la plupart des QoI par liaison hydrogène.
Par ailleurs, le résidu H181 de la sous-unité ISP, ligand du cluster fer-soufre, interagit par liaison hydrogène uniquement avec les QoI-D. Ceci entraîne un positionnement du domaine catalytique de l'ISP à proximité de l'hème bl du cytochrome b (position « b »). Un positionnement similaire s'observe en présence de famoxadone ou de fénamidone, mais ces deux fongicides n'ont pas d'interaction directe avec le résidu H181 de l'ISP.
Enfin, les autres QoI-P entraînent un positionnement du domaine catalytique de l'ISP à proximité de l'hème du cytochrome c1 (position « c1 ») (Berry and Huang 2011 ; Esser et al., 2004)(Tableau 1).
Mutations du gène codant le cytochrome b
Résistances aux QiI et QioI en laboratoire
De nombreux mutants faiblement résistants à l'antimycine A, à l'ilicicoline H, au diuron, au HDQ ou au HQNO (QiI), ainsi qu'à la funiculosine (QioI) ont été induits au laboratoire, notamment chez S. cerevisiae. Les changements concernent des résidus du site Qi, compris entre les positions 15 et 40 (ex. : Q22E/T, G33A) ou les positions 195 à 230 (ex. : L198F, H204Y, M221Q, F225L, K228M).
L'amétoctradine présente une légère activité sur la levure, mais aucune des substitutions précédentes n'affecte sa toxicité. Par contre, les substitutions N208V et R218K induisent une sensibilité réduite, suggérant que cette triazolopyrimidine pourrait secondairement se fixer sur le site Qi.
Dans les mêmes conditions expérimentales, l'amisulbron et la cyazofamide n'ont aucune activité sur S. cerevisiae (Fisher and Meunier 2008 ; Brasseur et al., 1996). Pour l'instant, aucune résistance acquise de cible à ces divers fongicides n'a été signalée chez des oomycètes.
Étant donné que l'amétoctradine et les diméthyl-sulfonamides sont peu ou pas actifs sur les eumycètes, il est probable que leur résistance naturelle résulte de différences structurales, respectivement des sites Qo et Qi, liées à des résidus particuliers.
Les comparaisons des séquences du cytochrome b entre les eumycètes et les oomycètes dans les régions mentionnées précédemment pourraient permettre de trouver des résidus candidats pour des mutations chez les oomycètes.
Toutefois, une résistance croisée entre ces deux types d'inhibiteurs du complexe III, liée à un changement qualitatif du cytochrome b, semble improbable.
Résistances aux QoI en pratique : les substitutions G143A et F129L
Une forte résistance aux strobilurines est déterminée par la substitution G143A chez une trentaine de champignons phytopathogènes, en particulier des ascomycètes responsables d'oïdiums, de la tavelure du pommier et de la septoriose du blé, et de quelques oomycètes dont l'agent responsable du mildiou de la vigne Plasmopara viticola.
Ainsi, les souches de P. viticola sont résistantes à tous les QoI-P (y compris la famoxadone et la fénamidone) mais demeurent sensibles aux QoI-D et QiI.
Par ailleurs, certaines espèces fongiques sont à l'abri d'un tel changement car elles comportent un intron entre les codons 143 et 144, conduisant à des mutations létales - cas de nombreuses rouilles, certains Alternaria spp. et Monilia spp. et Pyrenophora teres, responsable de l'helminthosporiose de l'orge. Botrytis cinerea, agent de la pourriture grise, se singularise par la présence de souches avec ou sans introns ; seules ces dernières peuvent porter la substitution G143A (Fernandez-Ortuno et al., 2008 ; Leroux and Walker 2010b)(Tableau 2 page suivante).
La substitution F129L, observée chez quelques champignons phytopathogènes dont Alternaria solani, Pythium aphanidermatum, P. teres et P. viticola, détermine une résistance faible à modérée aux QoI-P et à la stigmatelline mais pas à l'atovaquone (Fisher and Meunier 2005). Il conviendrait de vérifier le comportement de l'amétoctradine vis-à-vis de souches de P. aphanidermatum et de P. viticola présentant cette substitution qui concerne un résidu de l'hélice transmembranaire C du cytochrome b (Tableau 2).
Substitutions G137R/S, M139I/L, I147T/V, S152T/A/F
Chez Pyrenophora tritici-repentis, agent de l'helminthosporiose du blé, la substitution G137R entraîne une résistance faible à modérée aux strobilurines.
La substitution G137S n'affecte pas l'activité de l'acéquinocyl chez l'acarien T. urticae, mais détermine une faible résistance au bifénazate (Tableau 2).
Une résistance modérée à l'atovaquone est observée avec les substitutions M139I/L, respectivement chez les protozoaires responsables du paludisme et de la toxoplasmose.
Chez T. urticae, la substitution I147T détermine une résistance uniquement au bifénazate alors que chez une souche sélectionnée au laboratoire, la substitution S152F entraîne une résistance croisée entre le bifénazate et l'acéquinocyl ; ces deux substitutions sont systématiquement combinées avec le changement G137S.
Une résistance faible à modérée vis-à-vis de l'atovaquone est observée chez P. jiroveci, avec les substitutions I147V ou S152A. Lorsque celles-ci sont exprimées dans S. cerevisiae, le premier changement détermine une résistance à d'autres QoI-D dont l'amétoctradine et la stigmatelline, ainsi qu'à l'azoxystrobine ; en revanche, la sensibilité reste inchangée à ces trois QoI pour le second changement. Ces différents acides aminés sont localisés dans la boucle cd, comme le résidu G143 (Tableau 2).
Quelques autres substitutions
Les substitutions P266L, I269L/M ou L275F confèrent une faible résistance à l'atovaquone chez les agents responsables du paludisme, de la pneumocystose ou de la toxoplasmose.
Dans le domaine PEWY, la substitution P271T détermine une résistance moyenne à forte vis-à-vis de l'acéquinocyl et du bifénazate chez T. urticae.
Enfin, les substitutions F278L, Y279N/S/C et L282V/F confèrent une forte résistance à l'atovaquone chez des agents du paludisme. Exprimés chez S. cerevisiae, ces changements déterminent aussi une résistance à des strobilurines mais pas forcément à la stigmatelline (Brasseur et al., 1996 ; Fisher & Meunier 2005, 2008 ; Van Nieuwenhuyse et al., 2009) (Tableau 2).
Quelle hérédité pour ces résistances ?
En cas de changement qualitatif du cytochrome b, la résistance aux inhibiteurs du complexe III aura une hérédité monoparentale (généralement maternelle) car le gène impliqué est mitochondrial.
Chez certains champignons phytopathogènes et T. urticae, la résistance respectivement à des QoI-P et des QoI-D est parfois instable du fait d'une hétéroplasmie (présence dans la même cellule de mitochondries normales et mutées).
Toutefois, dans de nombreuses situations, la résistance apparaît homoplasmique (présence uniquement de mitochondries mutées) ; la sélection opère alors comme pour une résistance chromosomique ou nucléaire, observée pour la plupart des fongicides unisites (Fernandez-Ortuno et al., 2008 ; Van Leeuwen et al., 2010).
Quelle « fitness » chez les résistants ?
Les changements au niveau du cytochrome b sont susceptibles d'affecter le fonctionnement du complexe III et de ce fait, la fitness (capacité de survie et de compétition) des souches résistantes.
Ainsi, les substitutions F129L, G143A et F278I ont des effets mineurs alors que les substitutions S152A, Y279C/S et L282V affectent l'activité catalytique.
Chez Plasmodium falciparum, une surexpression de gènes codant pour d'autres sous-unités du complexe III pourraient compenser l'effet délétère de la substitution Y279S.
Dans le cas de la substitution G143A, des différences de fitness ont été reportées entre espèces fongiques (Fisher and Meunier 2008).
Résistance non liée au cytochrome b, mécanismes divers
L'AOX entre en scène
Une résistance simultanée à l'ensemble des inhibiteurs du complexe III ainsi qu'à ceux de complexe IV (ex. : cyanure) peut résulter du fonctionnement d'une alternative oxydase (AOX) ou ubiquinol oxydase. Chez certaines espèces fongiques comme Botrytis cinerea et Fusarium graminearum, la faible efficacité en pratique des strobilurines résulterait de cette respiration alternative qu'est l'AOX. Une résistance acquise liée à une expression accrue de l'AOX a été montrée chez Venturia inaequalis (essentiellement au niveau des conidies) et plus récemment chez P. viticola. Dans ce dernier cas, la résistance croisée positive entre l'amétoctradine et les diméthyl-sulfonamides a été clairement démontrée.
De façon générale, l'impact sur l'efficacité pratique des inhibiteurs du complexe III demeure inconnu, du fait de fréquences faibles et probablement d'une fitness réduite (à vérifier chez P. viticola).
L'AOX pourrait cependant permettre la survie (à des doses sublétales d'inhibiteurs du complexe III) de souches de bioagresseurs susceptibles ensuite de développer de fortes résistances liées à des mutations du gène codant le cytochrome b (Wood & Hollomon 2003).
Par ailleurs, chez Phytophthora capsici (en présence de SHAM, un inhibiteur de l'AOX), des souches fortement résistantes à la cyazofamide ont été détectées aux États-Unis ; le mécanisme en cause reste inconnu (Kousik and Keinath 2008).
Mutations hypothétiques dans un gène nucléaire
Le site Qo étant en contact avec la sous-unité ISP, il est possible qu'un changement structural de cette protéine puisse déterminer une résistance aux QoI-P et/ou QoI-D.
À ce jour, aucun cas de résistance à des QoI, lié à une mutation dans le gène nucléaire codant l'ISP n'a été démontré. Mais cette possibilité a été évoquée pour des populations de T. urticae et de Podosphaera fusca (agent responsable de l'oïdium des cucurbitacées) moyennement à fortement résistantes à des strobilurines sans présenter de mutation au niveau du cytochrome b (Fernandez-Ortuno et al., 2008 ; Van Nieuwenhuyse et al., 2009).
La MDR, « multidrug resistance » : rôle des transporteurs membranaires
La production accrue de transporteurs membranaires de type ABC ou MFS, susceptibles d'excréter une grande variété de substances toxiques présentant des modes d'action différents, peut conduire à la « multidrug resistance » (MDR).
Ce phénomène, observé chez des souches de B. cinerea et de M. graminicola collectées au champ, concerne des strobilurines et l'antimycine A.
De façon générale, les niveaux de résistance sont faibles. Dans le domaine agricole, ce mécanisme à lui seul a peu de chance d'entraîner des baisses notables de l'efficacité des inhibiteurs du complexe III.
En revanche et comme dans le cas de l'AOX, les souches MDR pourraient mieux survivre aux traitements, puis développer des résistances fortes liées à des mutations dans le gène codant le cytochrome b. Ce phénomène a été décrit pour diverses substances antipaludiques mais pas l'atovaquone, et n'a pas été rapporté chez les acariens et les oomycètes (Leroux & Walker 2010b ; Rosenthal 2013).
Que penser de la détoxification ?
Un mécanisme fréquemment rencontré pour des acaricides, insecticides et herbicides est la détoxification par l'intermédiaire de glutathion-S-transférases, de monooxygénases à cytochrome P450, d'estérases ou d'hydrolases.
Ce phénomène peut, comme la MDR, conduire à une résistance croisée entre pesticides à modes d'action différents. Toutefois, les observations réalisées à ce jour indiquent que la détoxification est en réalité un mécanisme mineur de résistances aux inhibiteurs du complexe III.
Pour l'anecdote, des souches de V. inaequalis résistantes au krésoxim-méthyl détoxifient cette strobilurine via une estérase. Par ailleurs, dans le cas de T. urticae, un ou des cytochromes P450 seraient impliqués dans la résistance à des inhibiteurs des complexes I ou II (Van Leeuwen et al., 2010).
Comment gérer ces résistances ?
Conséquences pratiques variables
Nous l'avons vu plus haut, les inhibiteurs respiratoires du complexe III ayant des utilisations majeures en médecine humaine et dans le domaine phytosanitaire peuvent être classés en trois catégories : QoI-P, QoI-D et QiI (Tableau 1).
Nous avons montré qu'ils pouvaient tous être concernés par la résistance, principalement à cause de mutations affectant leur cible commune, le cytochrome b (Tableau 2).
Une partie des mutations décrites plus haut a été détectée chez des isolats naturels et a conduit à des pertes d'efficacité pratique.
Cependant, au cas par cas de chaque mutation, les spectres et niveaux de résistance, et donc les conséquences pratiques, peuvent être variables. Dans ce contexte, il est primordial de mettre en oeuvre des stratégies antirésistance le plus précocement possible.
Gestion des fongicides QoI-P
Parmi ces inhibiteurs respiratoires, les plus nombreux sont les QoI-P de type strobilurines avec leur large spectre d'activité antifongique. De nombreux ascomycètes et oomycètes phytopathogènes sont touchés par la résistance à cette classe d'inhibiteurs du complexe III, mais quelques espèces majeures échappent encore à ce phénomène, comme les agents des rouilles, le mildiou de la pomme de terre ou la pyriculariose du riz.
En terme de stratégie antirésistance, des limitations d'utilisation des fongicides QoI-P, allant jusqu'à l'abandon, sont préconisées.
Les associations avec d'autres modes d'action doivent comporter une dose suffisante du produit compagnon, lui-même non concerné par des risques de résistance, pour qu'une efficacité correcte soit assurée même en cas de résistance généralisée.
Parmi les fongicides associés aux QoI-P, on trouve des multisites (ex. : antimildiou) et aussi des unisites, notamment des SDHI. Ce dernier type d'association est toutefois menacé par la sélection de souches résistant simultanément aux QoI et aux SDHI (ex. : B. cinerea ; Alternaria sp.).
Gestion des antipaludiques et des acaricides QoI-D
Pour la lutte contre le paludisme, la stratégie retenue est d'associer l'atovaquone avec un antifolate, le proguanil. De plus, des recherches sont conduites pour mettre au point d'autres inhibiteurs du complexe III efficaces sur les mutants de Plasmodium les plus fréquents, notamment ceux portant la substitution Y279S (Rosenthal 2013).
Concernant l'agriculture, une limitation à une ou deux applications par saison et une alternance avec d'autres modes d'action sont préconisés pour améliorer la durabilité des acaricides acéquinocyl et bifénazate (Van Nieuwenhuyse et al., 2009).
Gestion de l'amétoctradine
Le cas de l'amétoctradine et des diméthyl-sulfonamides (anti-oomycètes spécifiques respectivement QoI-D et QiI) apparaît singulier.
En effet, la résistance liée à une modification de leur cible commune n'a jamais été décelée au champ. De plus, il est improbable qu'une mutation affectant soit le site Qo, soit le site Qi entraîne une résistance croisée forte entre ces deux types d'inhibiteurs du complexe III.
En revanche, une résistance croisée a été observée chez des souches de P. viticola surexprimant une alternative oxydase (AOX).
Par précaution, une limitation de l'emploi de ces deux classes d'anti-oomycètes s'impose avec, sur vigne, un maximum global de trois traitements par saison et de deux par type d'inhibiteur. Leur association avec un mode d'action non concerné par la résistance (ex. : multisite, phosphonate) apporte une sécurité supplémentaire pour gérer la résistance AOX mais aussi pour prévenir les résistances spécifiques à ces deux catégories d'inhibiteurs.
La surveillance des populations de P. viticola est indispensable pour détecter des souches moyennement à fortement résistantes à ces fongicides. Elle doit être conduite indépendamment avec ces deux types d'inhibiteurs du complexe III.
Rappelons que la présence généralisée de souches fortement résistantes aux seuls QoI-P (substitution G143A du cytochrome b) limite l'intérêt de ce type d'inhibiteurs du complexe III dans la lutte contre le mildiou de la vigne (voir la note commune Maladies de la vigne 2015).
SUMMARY
ABSTRACT - Respiratory inhibitors of the mitochondrial complex III are used against phytopathogenic fungi, acari and human infections.
They affect the binding of the coenzyme Q to the cytochrome b, one of the major components of complex III.
They are classified into three mains groups : QiI, QoI-P and QoI-D.
Resistance to these inhibitors is related to the alteration of their common target site of action but other mechanisms may occur.
Therefore, restricted use and combinations or alternation with other modes of action are needed.
KEYWORDS - Mitochondrial respiration complex III, cytochrome b, QiI, QoI-D, QoI-P, fungicide, acaricide, ametoctradine, resistance.
Fig. 1 : Cibles des principaux produits phytosanitaires affectant spécifiquement la respiration
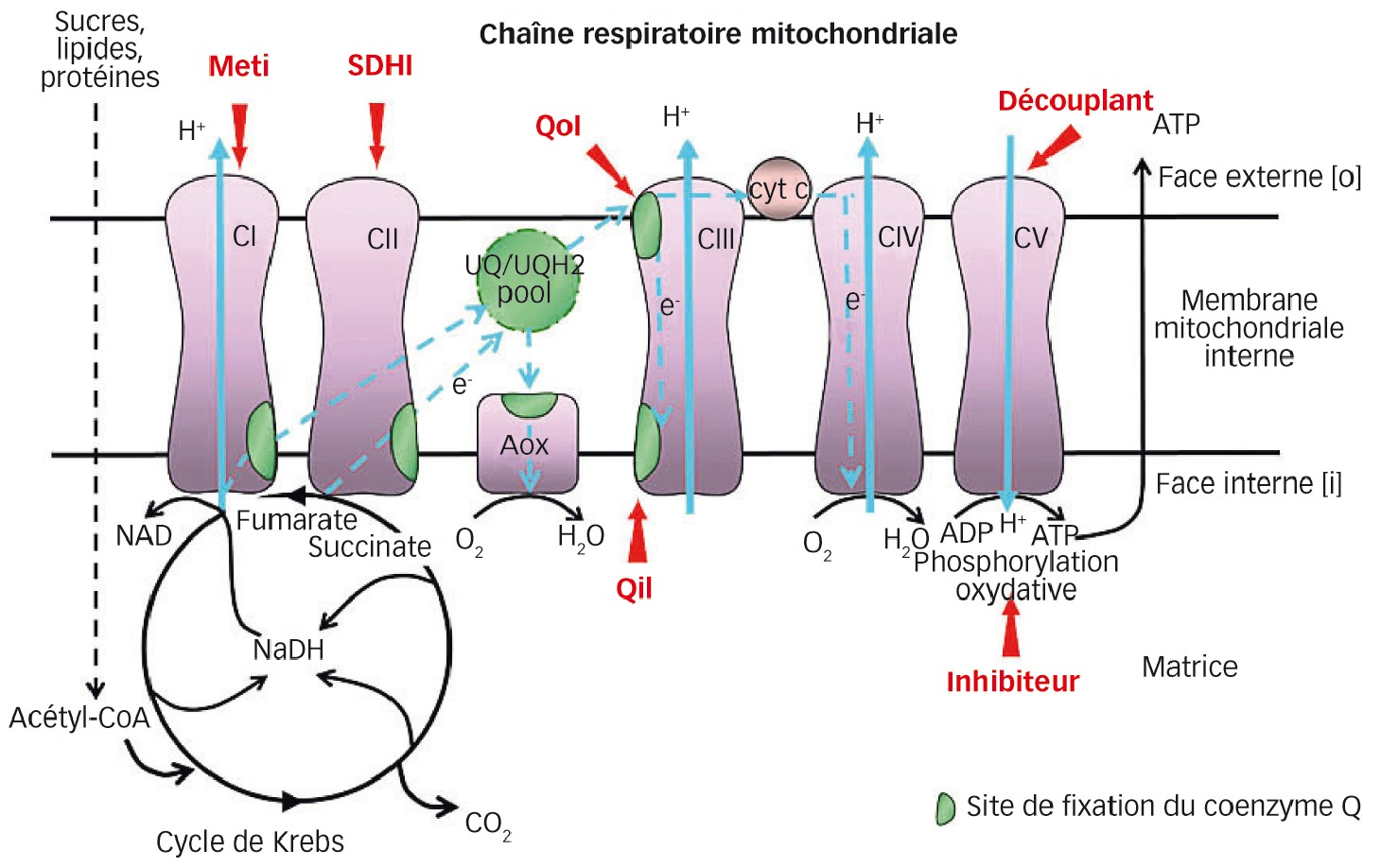
En rouge, les modes d'action désignés par leurs abréviations utilisées en phytopharmacie.
Fig. 2 : Structure partielle du complexe mitochondrial III ou bc1 et inhibiteurs
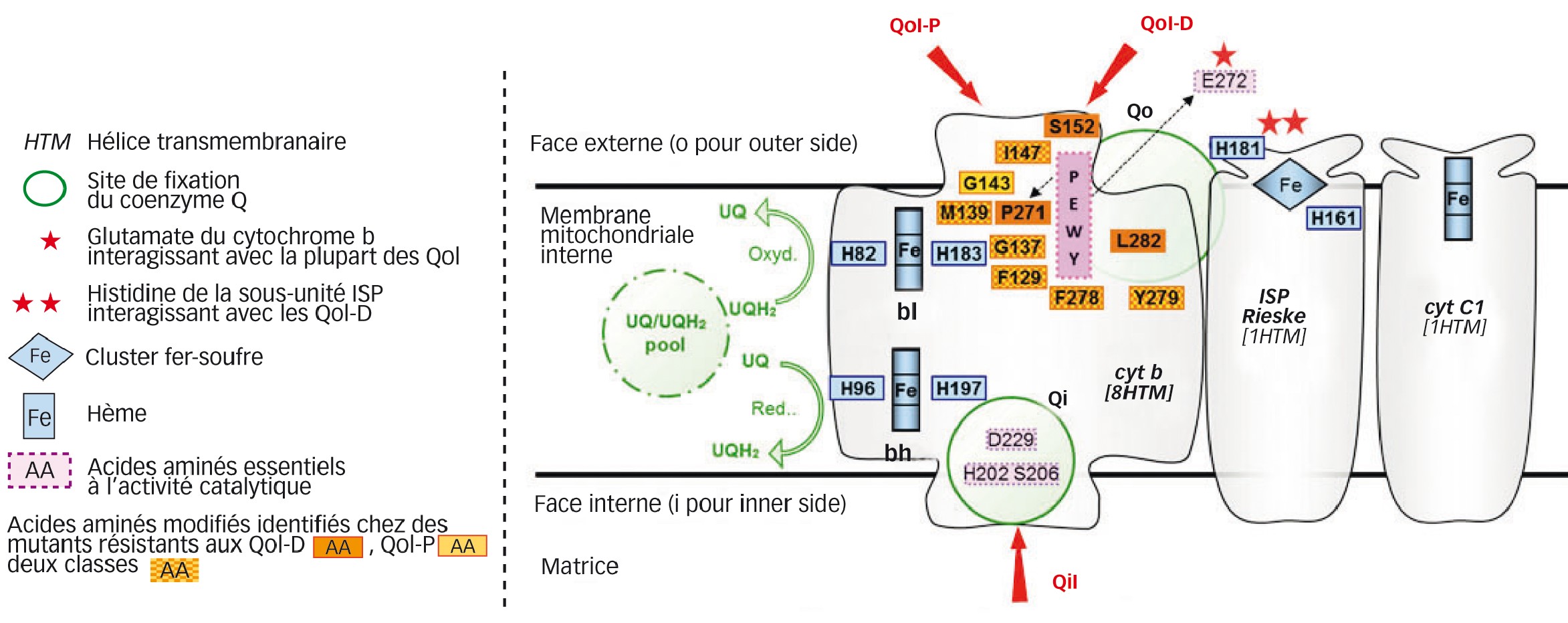
« Détail » de la Figure 1 au niveau du « CIII ». On retrouve les QiI (en bas) et les QoI (en haut), ces derniers différenciés entre QoI-P (P pour « proche » de l'hème bl) et QoI-D (D pour « distant » de l'hème bl).
L'amétoctradine, premier fongicide phytopharmaceutique QoI-D
Fongicide antimildiou récemment mis sur le marché et dont le mode d'action vient d'être élucidé, l'amétoctradine est un inhibiteur respiratoire agissant au niveau du complexe III (cytochrome b), tout comme les strobilurines (QoI) ou encore l'amisulbrom et la cyazofamide (QiI).
L'amétoctradine se fixe sur le site Qo du cytochrome b, dans une position similaire à celle de la stigmatelline (antibiotique) et de certains acaricides et médicaments.
Dans la littérature scientifique, toutes ces molécules sont classifiées comme QoI-D, car leur fixation au sein du site Qo est distante d'un des hèmes du cytochrome b, alors que celle des strobilurines (QoI-P) en est proche.
Le Frac a proposé la nomenclature spécifique QoSI pour l'amétoctradine.
Au final, il faut donc compter trois modes d'action indépendants, et ne présentant donc pas de résistance croisée entre eux, ciblant le cytochrome b : les QoI-D (ou QoSI) et QoI-P (site Qo) et les QiI (site Qi).
Des souches présentant une résistance croisée entre l'amétoctradine et les QiI ont récemment été détectées à faible fréquence au vignoble. Cette résistance non spécifique est causée par la surexpression de l'alternative oxydase (AOX), une enzyme permettant de compenser l'inhibition du cytochrome b et de produire l'énergie cellulaire par une voie alternative. La survie et la compétitivité (fitness) de tels isolats sont souvent compromises chez d'autres espèces fongiques, même si cela n'a pas été démontré formellement chez P. viticola.
À notre connaissance, les populations françaises de P. viticola ne comportent pas d'isolats résistant spécifiquement à l'amétoctradine.
Mais, comme pour tout unisite, une prévention appropriée de la résistance est à mettre en place précocement, car des mutants naturels résistant spécifiquement aux QoI-D après modification de leur cible sont détectés chez des acariens phytophages et les agents du paludisme et de la pneumocystose.
Note commune résistance vigne
La note Maladies de la vigne 2015 a été élaborée par un groupe de travail DGAL-SdQPV/Anses-RPP/Inra/CIVC/IFV/chambres d'agriculture. Elle fait le point sur les résistances du mildiou, de l'oïdium et du botrytis alias pourriture grise sur la vigne à l'issue de la campagne 2014. Elle donne les conseils qui en découlent en 2015. Pour la trouver, voir « Liens utiles » ci-dessous.

