 Imprimer
Imprimer
Laitues en pot utilisées pour les tests de sensibilité aux pourritures du collet. Les essais ont été menés sur 53 variétés. Photo : Inra

Feuilles détachées de laitue 48 heures après inoculation de mycelium de S. sclerotiorum. Photo : Inra
Toute nouvelle variété de laitue se voit imposer, en préalable à son inscription au catalogue officiel et son utilisation en production, l'épreuve DHS (distinction, homogénéité, stabilité). Cette épreuve contient des tests de résistance aux maladies mais, parmi celles-ci, les agents de pourriture du collet ne sont pas prioritaires. Quelles sont les conséquences sur le niveau de résistance des variétés actuelles ?
Comment sont gérées les maladies de la laitue
Sélection : pourritures en quatrième classe
De façon générale, les couples hôte/bioagresseurs sont regroupés en quatre classes :
- classe 1, avec obligation de test de résistance et déclaration systématique de l'obtenteur de la variété ;
- classe 2, avec test obligatoire uniquement si la variété est déclarée résistante ;
- classe 3, avec test chez l'obtenteur permettant de qualifier une variété vis-à-vis d'un bioagresseur appartenant à cette classe ;
- classe 4, dont les problèmes phytosanitaires sont reconnus mais non pris en charge durant le processus d'inscription des variétés ; c'est le cas pour Botrytis cinerea et Sclerotinia sclerotiorum sur laitue.
Du fait de cette réglementation et en l'absence d'évaluation de la valeur agronomique, technologique et environnementale (Vate), la résistance aux agents de la pourriture du collet contenue dans les variétés cultivées de laitue n'est pas connue et ne fait pas l'objet d'une communication de la part des obtenteurs de variétés. En revanche, lors du processus de sélection variétale, les sélectionneurs de laitue observent depuis des décennies le comportement des plantes en culture vis-à-vis de la pourriture du collet. Il pourrait donc exister un processus de sélection indirect écartant les accessions les plus sensibles.
Usage actuel des fongicides en lien avec les autres pratiques
La gestion du mildiou et des champignons nécrotrophes en agriculture conventionnelle repose largement sur des applications de pesticides et sur des mesures prophylactiques. Pour le mildiou, la protection chimique vise à diminuer la pression de sélection sur des souches de Bremia qui contourneraient les résistances variétales. Face aux agents de la pourriture du collet, la protection chimique (mêmes substances actives appliquées contre B. cinerea et Sclerotinia spp.) est le principal mode de lutte, accompagnant la lutte biologique et des mesures prophylactiques.
Les délais avant récolte (DAR) légaux des fongicides sont souvent longs au regard de la durée du cycle de culture de la laitue, comprise entre 60 et 90 jours. Ces DAR se situent en général entre 14 et 28 jours. De ce fait, et parce que la plupart des matières actives n'ont pas d'action curative, les fongicides sont surtout appliqués de façon préventive (Barrière et al., 2014).
En culture d'hiver, période la plus propice au développement des champignons, en moyenne trois à quatre traitements contre le mildiou et trois traitements contre les agents de la pourriture du collet sont appliqués (Barrière et al., 2015). La résistance des agents pathogènes à diverses classes de fongicides est bien documentée (par exemple, Lamichlane et al., 2016), et le nombre de traitements est en partie motivé par la nécessité de traiter avec des pesticides de différentes classes afin de minimiser les épidémies causées par des souches résistantes aux fongicides.
Pourquoi la sélection ne vise pas plus directement les pourritures du collet
Le développement de variétés présentant une sensibilité réduite à B. cinerea et Sclerotinia spp. pourrait permettre de limiter la lutte chimique, mais l'introduction, par des techniques classiques de sélection, de résistances partielles de nature polygénique exige un effort long, difficile et coûteux. Parmi les divers caractères sélectionnés, la demande continue de nouvelles résistances aux souches de Bremia lactucae conduit à un turn-over très rapide des variétés.
Chaque année, plusieurs dizaines de variétés nouvelles sont inscrites au catalogue officiel et leur durée de vie moyenne n'excède guère deux ou trois saisons. On peut se demander si, au cours du processus de sélection, le retrait des génotypes les plus sensibles suffit pour obtenir des niveaux significatifs de résistance à la pourriture du collet.
Quel effet, sur ces pourritures, de la sélection visant d'autres bioagresseurs ?
Dans ce contexte, nous avons cherché à savoir si les variétés actuelles du catalogue présentaient une sensibilité particulière à B. cinerea et à S. sclerotiorum, en comparaison avec des laitues inscrites il y a plusieurs décennies et tombées dans le domaine public. À des fins de simplification, le travail est focalisé sur la batavia, principal morphotype cultivé en volume en France (voir Encadré 1).
La sensibilité est ici désignée comme l'intensité des symptômes observés dans des tests d'infection en conditions maîtrisées au laboratoire, sur feuilles détachées. Représentant l'expression d'une résistance physiologique des plantes, elle ne permet pas totalement de présager de leur comportement au champ. Ce dernier dépend, outre cette résistance physiologique, de caractéristiques architecturales des plantes (Hayes et al., 2010). Mais des résultats d'essais préliminaires dans nos stations expérimentales montrent une corrélation significative entre la sensibilité observée au laboratoire et celle observée au champ après inoculations artificielles.
Le terme résistance physiologique utilisé ici correspond à l'expression physiologique d'une résistance d'origine génétique, susceptible d'être influencée par les conditions de l'environnement. Dans la mesure où les caractéristiques architecturales des plantes sont également liées au génotype et qu'elles peuvent être à l'origine d'une moindre sensibilité à la pourriture du collet, la résistance physiologique ne recouvre qu'une partie de la résistance génétique.
Une évaluation au laboratoire
Cinquante-trois variétés
Quatre essais de phénotypage ont été menés au printemps (deux essais) et à l'automne (deux essais) 2015. Cinquante-trois variétés - en quasi-totalité des batavias - ont été testées, dont trois variétés anciennes du domaine public et cinquante inscrites au catalogue issues de sept obtenteurs différents (entre deux et quatorze variétés par obtenteur), plus des deux tiers inscrites après 2007 et une seule avant 2000. Pour des raisons de confidentialité, les noms des variétés et des obtenteurs ne sont pas donnés ici.
Quarante-trois variétés ont été testées dans deux essais, et dix variétés constituant un panel de référence dans les quatre essais. Dans chaque essai, trente variétés étaient testées avec dix plantes par variété : cinq plantes utilisées pour tester la sensibilité à B. cinerea, et cinq pour tester la sensibilité à S. sclerotiorum.
Tests sur feuilles détachées en survie
La méthodologie complète des tests a été décrite par Barrière (2015). Elle est rappelée ici brièvement. Les plantes ont été repiquées avec un terreau commercial dans des pots de 1,5 l quelques jours après le semis. Les tests ont été réalisés sur feuilles détachées sur des plants d'environ six semaines.
Sur chaque plante, deux feuilles de la couronne intermédiaire, d'environ 12 cm de longueur, ont été détachées et placées dans des boîtes de plexiglas sur un papier de ouate imbibé d'eau. Un disque de mycélium de 3 mm de diamètre a été déposé au centre de chaque feuille.
Le mycelium a été prélevé avec un emporte-pièce dans une boîte de Petri contenant un milieu de culture (Potato Dextrose Agar) ensemencé avec le champignon 72 heures auparavant. Deux souches différentes de chaque agent pathogène ont été utilisées : BC1 et SAS405 pour B. cinerea, SS1 et SS2 pour S. sclerotiorum. Les deux feuilles disposées dans les boîtes ont été inoculées chacune avec une souche.
Les boîtes ont ensuite été déposées dans des chambres de culture à température et humidité constantes. Les surfaces de lésion ont été mesurées, à l'aide d'un logiciel d'analyse d'image, après 48 heures pour S. sclerotiorum et 72 heures pour B. cinerea, car le développement de Sclerotinia est plus rapide.
Au total, 2 400 mesures de lésions ont été effectuées : trente variétés par essai, cinq plantes par variété, quatre souches de champignon et quatre essais. Des feuilles avec des lésions de S. sclerotiorum sont visibles sur la photo ci-dessus.
Mode de calcul pour classer les variétés
Afin d'intégrer les résultats des quatre essais, un indicateur robuste a été utilisé : le classement par rang de sensibilité des plantes. Dans chaque essai, pour chaque champignon et chaque souche, les surfaces de lésion mesurées sur chaque plante ont été classées par ordre croissant, la plante ayant la surface de lésion la plus faible recevant le rang 1, et celle ayant le niveau de lésion le plus élevé le rang 150.
Le rang moyen d'une variété correspond à la moyenne des rangs obtenus dans chaque essai, soit la somme des dix rangs mesurés divisée par dix pour les variétés testées dans deux essais, et la somme des vingt rangs divisée par vingt pour les dix variétés testées dans les quatre essais. Nous avons utilisé ce rang moyen pour présenter les résultats.
Résultats
La différence vient à 90 % de la variété
Nous avons tout d'abord étudié les différences de surfaces de lésions mesurées entre les essais et entre les souches (Figure 1).
Pour Botrytis, une grande homogénéité de surface de lésion a été enregistrée d'un essai à l'autre, mais des différences d'agressivité de souches - ou de résistance des hôtes - sont apparues : SAS 405 a systématiquement provoqué des dégâts inférieurs à BC1.
Pour S. sclerotiorum, les tests statistiques n'ont fait apparaître ni effet souche, ni effet essai, mais, globalement, il a été enregistré une variabilité plus forte de tailles de lésions que pour B. cinerea. À souligner : une part de la variabilité observée a pu être due à la combinaison différente de variétés utilisées à chaque essai ; ceci justifie le choix, comme indicateur, du rang de classement qui intègre cette variabilité.
Cependant, en moyenne pour une souche, la variance de la taille des lésions dans un essai représentait 87 % de la variance totale observée sur les quatre essais (les valeurs allaient de 52 % à 130 %). Cela indique que près de 9/10e de la variabilité des tailles de lésions provenait de différences entre les variétés d'un même essai, et que seule une faible part de la variabilité provenait de différences entre essais.
Les différences variétales dépendent peu des souches des pathogènes
Dans un second temps, nous avons observé les différences variétales de sensibilité aux deux champignons. Pour l'ensemble des variétés testées, la corrélation entre la sensibilité à BC1 et SAS405 est de 31 %, celle entre SS1 et SS2 de 61 %. Ceci indique que les deux souches de Botrytis ont généré des classements de sensibilité des variétés un peu différents, tandis que pour Sclerotinia les différentes souches ont faiblement affecté le classement de sensibilité des variétés. Ces différences entre souches n'affectant pas les principales conclusions obtenues, nous présentons, pour illustrer les résultats, les classements globaux par champignon sans distinction de souches.
Les variétés ont été regroupées par obtenteur (nommés dans les graphiques « Selec 1 » à « Selec 7 », en distinguant les variétés anciennes issues du domaine public « Public »), et le niveau de classement moyen de chaque groupe, pour l'ensemble de ses variétés, est donné Figure 2.
Des variétés modernes un peu plus sensibles à Botrytis et nettement plus à Sclerotinia
Pour B. cinerea, les tests statistiques montrent une différence significative (p = 0,04 : il y a quatre chances sur cent pour que cette différence soit due au hasard) mais assez faible, entre les variétés. Le regroupement par obtenteur ou avec les variétés du domaine public montre que ces dernières sont légèrement moins sensibles que les variétés modernes, mais la différence n'est significative qu'avec celles de l'obtenteur 4 chez lequel deux variétés seulement ont été testées. D'autre part, la sensibilité la plus basse est celle d'une accession moderne.
Pour S. sclerotiorum en revanche, le groupe des variétés du domaine public se distingue nettement des variétés modernes issues des différentes maisons de sélection (p = 0,003 : il y a seulement trois chances sur mille pour que la différence soit due au hasard). Le niveau de sensibilité des variétés modernes des sept obtenteurs est remarquablement homogène. Cependant, deux variétés du sélectionneur 1 ont une sensibilité faible. Deux parmi les trois variétés du domaine public sont très peu sensibles à S. sclerotiorum, et sont les moins sensibles de l'ensemble du panel. La troisième a montré une sensibilité moyenne, classée dans le premier tiers du panel.
Chez ces deux variétés anciennes peu sensibles à S. sclerotiorum, l'une est la moins sensible à Botrytis (classée 1re contre BC1 et 6e contre SAS405), et l'autre est relativement peu sensible (classée 8e contre BC1 et 15e contre SAS405). Globalement, la corrélation entre sensibilité à B. cinerea et S. sclerotiorum est apparue assez faible, entre 0 et 20 % selon les souches comparées deux à deux. Ainsi, il est apparu au sein des accessions anciennes du domaine public des niveaux de faible sensibilité aux deux souches des deux champignons. Parmi les variétés modernes, très peu se sont rapprochées d'un tel comportement avec, de plus, des niveaux de sensibilité plus élevés à l'un et l'autre des champignons.
Conclusion
La sensibilité à ces pourritures a augmenté
Les tests menés dans nos laboratoires, en conditions contrôlées, sur la sensibilité de la laitue aux champignons de la pourriture du collet B. cinerea et S. sclerotiorum se sont révélés fiables et répétables, avec une variabilité inter-essais assez réduite. Ils ont permis de discerner des différences variétales de sensibilité, faibles pour Botrytis et fortes pour Sclerotinia.
En particulier, les variétés anciennes du domaine public se sont montrées nettement moins sensibles à S. sclerotiorum. Ceci suggère que la résistance physiologique contenue dans certaines variétés anciennes se serait érodée au cours du processus de sélection des variétés modernes, particulièrement intense dans le cas de la laitue. Une confirmation au champ de ces conclusions devra être obtenue avant d'en déduire des implications pour la sélection variétale.
Malgré la proximité phyllogénétique des deux espèces B. cinerea et S. sclerotiorum, il n'existe plus dans la plupart des variétés modernes de corrélation entre la sensibilité aux deux champignons, alors que les variétés anciennes les plus résistantes à Sclerotinia ont une bonne résistance à Botrytis.
« Remonter » les résistances est souhaitable
L'effort de sélection actuel en laitue porte en particulier sur la résistance au mildiou B. lactucae. Malgré cet effort, le contournement régulier des résistances par certaines souches de B. lactucae impose le maintien d'un usage soutenu de fongicides anti-oomycètes. La lutte contre les agents de la pourriture du collet repose actuellement largement sur les fongicides, mais on peut espérer que l'introduction de résistances quantitatives à ces agents, résistances potentiellement plus difficilement contournables, permettrait de diminuer la pression phytosanitaire.
La durabilité des systèmes de production de laitue repose donc en partie sur la sélection, peu perceptible d'après nos résultats, de variétés moins sensibles aux agents de la pourriture du collet.
Une sélection possible
Nous montrons ici qu'il existe, au sein de la variabilité génétique de L. sativa, des différences de résistance physiologique, dont il faudra confirmer qu'elles s'expriment au champ.
Ces résistances sont probablement à déterminisme polygénique. Présentes dans d'anciennes variétés cultivées, elles sont donc a priori plus rapides à introgresser dans les variétés modernes que des résistances issues de laitues sauvages.
La caractérisation de ces résistances aux niveaux génétique, moléculaire et physiologique exige des efforts de recherche (Mbengue et al., 2016). Ces travaux permettront d'aider les sélectionneurs à identifier et introduire dans les variétés modernes les caractères de résistance.
Un dialogue entre les laboratoires de recherche publique et privée est donc nécessaire pour proposer aux producteurs des variétés de laitues moins dépendantes des pesticides.
*Inra UR1115. **Inra UR1052. ***Inra UR407.
Fig. 1 : Peu de différences entre souches et entre essais
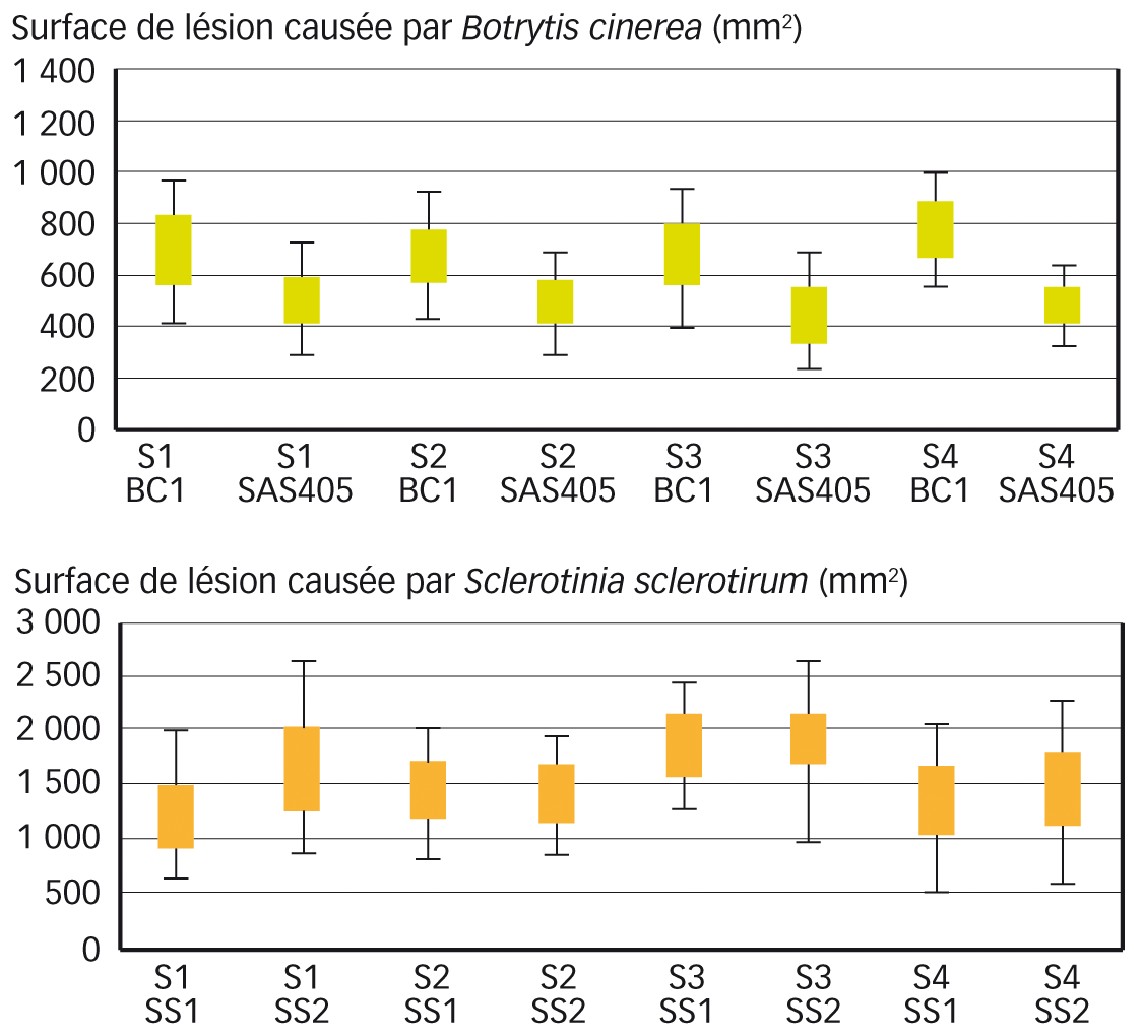
Comparaison des surfaces de lésion causées par Botrytis cinerea (en haut) et Sclerotinia sclerotiorum (en bas), pour quatre essais (S1 à S4) et deux souches de chaque champignon (BC1 et SAS405 pour B. cinerea, SS1 et SS2 pour S. sclerotiorum). Les symboles correspondent à des boîtes à moustaches : les rectangles définissent les valeurs comprises entre les 25e et 75e centiles de la distribution observée, la barre horizontale inférieure la limite du 5e centile, et la barre supérieure la limite du 95e centile : 90 % des valeurs mesurées sont situées entre les barres horizontales inférieures et supérieures.
Fig. 2 : Une évolution significative des sensibilités
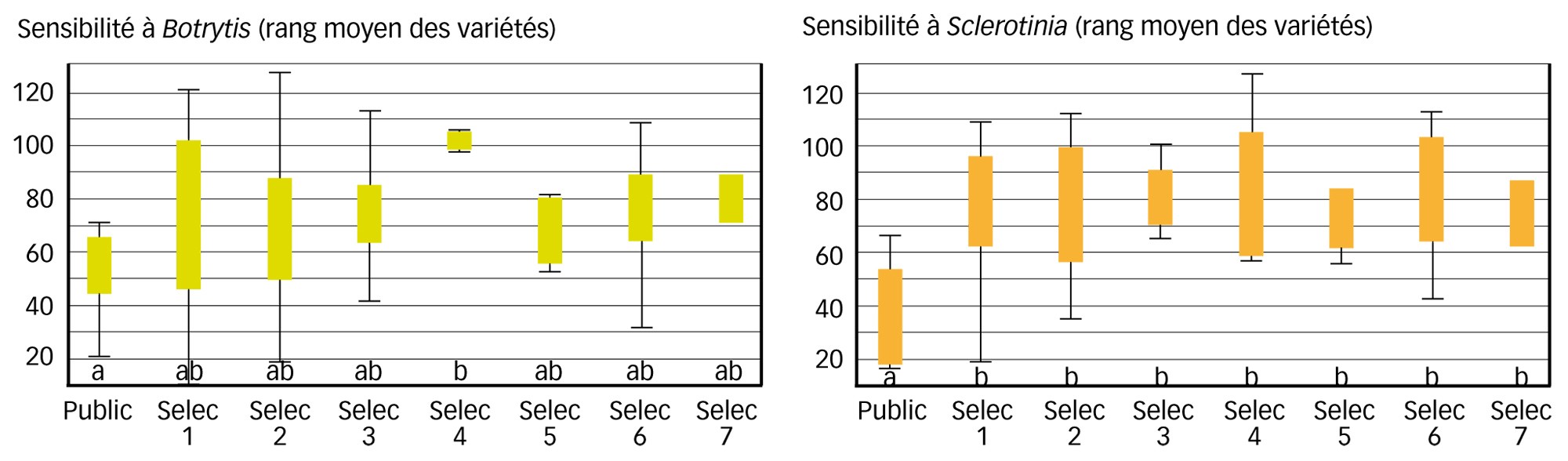
Sensibilité de 53 variétés de laitue à B. cinerea (à gauche) et à S. sclerotiorum (à droite). Données intégrant les résultats de quatre tests avec deux souches de chaque champignon par test. Variétés classées par niveau de sensibilité (moyenne des rangs par variété), et regroupées entre sept obtenteurs (« Selec 1 » à « Selec 7 ») ou en variétés du domaine public (« Public »). Un rang moins élevé indique une sensibilité plus faible des variétés. Les symboles correspondent à des boîtes à moustaches (voir légende de la Figure 1). Les lettres sous les symboles indiquent les différences significatives entre groupes, obtenues après analyse de variance (test de Student Newman-Keuls).
1 - Géographie, économie et bioagresseurs
Plante annuelle de la famille des Asteraceae, la laitue cultivée (Lactuca sativa L.) est produite toute l'année en plein champ ou sous abris dans des tunnels ou multichapelles, et pour une part croissante sous serre en hydroponie.
La laitue est produite et consommée dans le monde entier. Les principaux pays producteurs sont la Chine, les États-Unis, l'Inde, l'Iran et plusieurs pays européens dont l'Espagne, l'Italie et la France. Cette dernière produit environ 250 000 t annuellement(1). La production se segmente en divers morphotypes. En Europe, ce sont principalement les batavias (majoritaires en France), feuilles de chênes, laitues beurre, romaines et grasses.
Les bioagresseurs susceptibles de causer des dégâts sur les cultures de laitue sont nombreux et d'importance inégale selon les saisons : oomycètes ou champignons phytopathogènes (Bremia lactucae, Botrytis cinerea, Sclerotinia spp., Rhizoctonia solani, etc.), ravageurs (pucerons, noctuelles, thrips, nématodes, etc.), virus (LMV, le virus de la mosaïque de la laitue ; LRNA et MLBVV, agent des anneaux nécrotiques et virus des grosses nervures de la laitue...) ou bactéries (Xanthomonas campestris pv. vitians...).
(1) Sources : FAO, Eurostats.
2 - Maladies fongiques de la laitue et lutte génétique : panorama
Le bioagresseur le plus redouté parmi ceux s'attaquant aux cultures de laitue est l'oomycète Bremia lactucae (Regel), agent du mildiou de la laitue. Ce parasite obligatoire peut se conserver dans le sol sur des débris de culture ainsi que sur des Lactuca sauvages adventices. Il attaque les plantes à tous les stades et provoque des taches jaunes puis nécrotiques entre les nervures secondaires des feuilles (Blancard et al., 2003). Des sporanges dispersés par le vent et la pluie peuvent contaminer très rapidement des plantes adjacentes, propager l'épidémie et compromettre la récolte de parcelles entières.
Des résistances hôte-spécifiques basées sur l'existence de récepteurs à motif NBS-LRR (R gènes) sont déployées depuis plusieurs décennies dans les variétés à partir de gènes identifiés chez la laitue cultivée ou des espèces sauvages apparentées (Lebeda et al., 2014).
Cependant, l'évolution rapide du pathogène (Van Hese et al., 2016) exige d'introduire régulièrement de nouveaux gènes de résistances dans les variétés cultivées.
Les agents de la pourriture du collet, Botrytis cinerea, Sclerotinia sclerotiorum et Sclerotinia minor, sont des ascomycètes appartenant à la famille des Sclerotiniaceae.
Ces champignons nécrotrophes à très large spectre d'hôtes attaquent différents organes (feuilles, tiges, fleurs, fruits), détruisant leurs tissus pour en récupérer le contenu. Le cycle sexuel comprend la formation d'ascospores sur des fructifications issues des sclérotes qui sont les formes de conservation de ces champignons.
La contamination peut aussi avoir pour origine des hyphes mycéliens contenus dans des débris végétaux. Il existe également, chez B. cinerea, une multiplication végétative basée sur des conidies asexuées.
Une grande polymorphie génétique, associée à une production rapide et profuse d'inoculum et à de multiples facteurs de virulence, contribue à la sévérité des attaques de ces champignons (Van Kan, 2006 ; Bolton, 2006). La pourriture du collet provoque chaque année des pertes de rendement pouvant atteindre parfois 10 % de la culture.
Par contraste avec la lutte contre le mildiou, les sources de résistances génétiques à B. cinerea sont de nature hautement polygénique (Zang et al., 2016). Il n'a été observé contre B. cinerea, S. sclerotiorum et S. minor, quelle que soit la plante-hôte considérée, que des résistances partielles, aux mécanismes sous-jacents mal connus.
Chez la laitue, de telles résistances contre B. cinerea et Sclerotinia spp. ont été observées dans des accessions sauvages de L. virosa et L. saligna, (Whipps et al., 2002 ; Hayes et al., 2010 ; Shim et al., 2014).

