
 Imprimer
Imprimer")
Rouille brune inoculée (ph. B. Pariaud)
 des données sur des résistances spécifiques. ph. H. Goyeau")
Symptômes de rouille brune après inoculation dans un des essais au champ mis en place pour évaluer des résistances partielles, et qui permettent aussi de valider (ou non) des données sur des résistances spécifiques. ph. H. Goyeau
La protection fongicide contre les maladies du blé en végétation pèse sur le budget des agriculteurs : en 2007, 68 €/ha en moyenne. Et le plan Ecophyto pousse à réduire ces traitements.
L'un des moyens de cette réduction est d'utiliser des blés présentant une bonne tolérance variétale aux principales maladies foliaires, en particulier celles qui justifient en moyenne deux traitements : la septoriose et la rouille brune causée par Puccinia triticina. Cette dernière est présente chaque année quasiment sur tout le territoire français et sa nuisibilité peut dépasser 20 q/ha(1). Cependant, pour qu'une résistance variétale contre cette maladie contribue au « développement durable », elle doit être durable au premier sens du terme : il faut qu'elle persiste dans le temps. Mais comment estimer voire prévoir la durabilité d'une résistance ? Pas facile...
La résistance d'une variété peut reposer sur des gènes de résistance dits spécifiques ou non : dans le cas dit spécifique, elle s'exprime par une réponse « tout ou rien » (résistant/sensible). Ce type de résistance a été très utilisé vu son déterminisme génétique simple (monogénique) et son niveau (résistance totale).
Résistances spécifiques et partielles, les enjeux
Une dizaine de gènes Lr de résistance spécifique à la rouille brune sont présents dans les variétés de blés cultivées en France, seuls ou en combinaisons de deux ou trois gènes (Goyeau et al., 2006). Mais ce type de résistance est très souvent contourné, parfois très vite après 2 à 3 années de culture (Kolmer, 2001). En effet les populations du champignon sont diversifiées pour les virulences et certains pathotypes cumulent jusqu'à 13 virulences. Baser une stratégie sur des gènes de résistance spécifique exige de suivre les populations pathogènes.
Les résistances de type non spécifique, comme celles dites partielles (slow-rusting pour les rouilles), sont réputées plus durables que les résistances spécifiques (Kolmer, 1996). Elles s'expriment par une quantité réduite de maladie et reposent sur un petit nombre de gènes mineurs (Kolmer, 1996). Après 25 années de sélection pour ce type de résistance vis-à-vis des rouilles des céréales, le CIMMYT(2) considère que combiner quelques gènes de résistance partielle à effet additif permet d'obtenir une résistance durable (Singh et al., 2005). Les mécanismes impliqués sont une augmentation de la période de latence et une réduction de la quantité de spores produites et/ou de l'efficacité d'infection de ces spores.
La prise en compte de ce type de résistance dans les programmes d'amélioration variétale est donc recommandée. Mais elle pose plusieurs questions :
– on connaît très peu de choses sur la durabilité effective des résistances partielles. On sait par expérience que le niveau de résistance d'un cultivar varie dans le temps et qu'une telle résistance risque de s'éroder au fil des années (Mundt et al., 2002). Mais on ne sait pas à quelle vitesse ni comment une population pathogène s'adapte à une résistance partielle ;
– on ne sait rien sur la diversité des populations parasites en termes de réponse à une résistance partielle. Un niveau de spécificité a été montré pour certains QTL(3) de résistance (Manzanares et al., 2000 ; Pflieger et al., 1999), mais la variabilité des composantes de l'agressivité n'a jamais été décrite à l'échelle d'une population. Tous les individus ont-ils la même agressivité sur une variété à résistance partielle, ou existe-til une diversité susceptible d'éroder rapidement le niveau de résistance (Lannou, 2001) ?
Triple travail
Dans l'objectif d'évaluer l'efficacité et la durabilité des résistances génétiques des types spécifique et partiel, nous avons donc :
• intensifié un réseau pré-existant de suivi des virulences des populations pathogènes ;
• développé une méthodologie d'étude de la résistance partielle en conditions contrôlées ;
• étudié le niveau d'agressivité(4) comme caractère quantitatif dans les populations parasites. On suppose en effet que l'érosion d'une résistance partielle résulte d'une sélection pour l'agressivité chez le parasite.
Réseau de suivi des populations : du prélèvement à l'inoculation
Des feuilles portant des pustules de rouille brune sont prélevées dans les parcelles non traitées de réseaux d'essais : principalement sélectionneurs du Club 5 et du CETAC avec des coopératives associées à leurs réseaux, ainsi qu'Arvalis-Institut du Végétal, mais aussi SRAL, chambres d'agriculture, sociétés phytosanitaires. L'échantillonnage est ciblé sur les variétés les plus cultivées (Apache, Caphorn et Orvantis de 2006 à 2008, plus Aubusson et Sankara à partir de 2007) tout en gardant des variétés de référence suivies sur le long terme (Soissons, Isengrain, Trémie, Charger).
L'accès à ces réseaux d'essais permet une bonne couverture géographique du territoire avec 16 à 20 régions représentées selon les années. Pour chaque site de prélèvement et chaque variété, un isolement monopustule est réalisé, donnant un isolat par lieu et par variété. Tous les isolats sont conservés sous forme de spores en collection à – 80 °C.
Les isolats collectés sont inoculés au stade plantule en serre à une série de 27 lignées ou variétés composant la gamme d'hôtes différentiels. La formule de virulence/avirulence sur 18 lignées de cette gamme permet de déterminer le pathotype (= combinaison de virulences) des isolats. La nomenclature des pathotypes est explicitée dans Goyeau et al. (2006).
Mesure de la résistance partielle en conditions contrôlées
À l'issue des essais menés en 2005 et 2006, 7 variétés et lignées ayant exprimé au champ une gamme de niveaux de résistance partielle ont été sélectionnées pour quantifier en conditions contrôlées les composantes de cette résistance. L'expérimentation a été menée en serre à l'aide d'une méthodologie simplifiée dérivée de celle mise au point par Pariaud et al. (2009 b) : en 2007 avec le pathotype P3 et en 2008 avec les pathotypes P3 et P4. Le couple P1 x Soissons a été inclus comme référence (forte agressivité, voir Pariaud et al., 2009 b) chaque année.
Pour chacune des variétés ou lignées Andalou, Aciento, Morocco (témoin sensible), Trémie, Soissons, Apache, PBI et LD7, quinze plantes adultes ont été inoculées au stade floraison (méthodologie exposée dans notre communication à la CIMA de décembre 2009, v. « Bibliographie » p. 24). De suite après l'inoculation, les plantes ont été mises en chambre de rosée à 15 °C entre 14 et 18 heures, puis replacées dans la serre jusqu'à la fin de l'expérience.
L'efficacité d'infection a été définie comme le rapport entre un nombre de lésions sporulantes en fin de période de latence et la quantité de spores déposées. Les mesures de quantité de spores déposées sur feuille et de quantité de lésions sporulantes (v. méthodologie dans notre communication à la CIMA) ont été faites sur 10 plantes de chaque traitement (variété x pathotype).
La période de latence est définie par le temps au bout duquel 50 % des lésions sont apparues (T50, Shaner, 1980).
La sporulation a été définie comme le rapport entre la quantité de spores produite et la surface sporulant par feuille drapeau. La quantité de spores a été estimée en pesant les spores récoltées dans des sachets de cellophane ouverts placés autour des feuilles drapeaux entre 17 et 21 jours après inoculation (jai). La surface sporulante a été estimée à 21 jai : les feuilles ont été coupées et photographiées à l'aide d'un scanner. Les images ont été analysées avec le logiciel Optimas (programmes développés par P. Belluomo, EGC, INRA Grignon).
L'analyse exploratoire des données et la sortie des graphiques a été effectuée avec le logiciel S plus (© Insightful Corp.) Les analyses statistiques ont été réalisées avec le logiciel JMP5 (© SAS Institut inc., Cary, NC, USA). Une transformation logarithmique des variables a été opérée pour respecter les conditions de linéarité du modèle. Les facteurs testés ont été le pathotype et la variété. La densité de lésions à la fin de la période de latence a été testée en covariable dans toutes les analyses. Les autres facteurs pris en compte ont été : le stade de développement du talle à l'inoculation (épiaison ou floraison) et la présence ou l'absence de lésions d'oïdium sur la plante.
Mesure de l'agressivité dans les populations parasites
Notre étude s'est basée sur l'analyse de la structure des populations de P. triticina en France qui avait montré que sur la variété Soissons un pathotype (073100, codé P1 dans la présente étude) dominait largement (Goyeau et al., 2006). La question de l'adaptation différentielle a été abordée en deux étapes successives :
1) Le pathotype P1 est-il plus agressif sur Soissons que deux autres pathotypes, P2 et P3, virulents sur Soissons mais présents en fréquence plus faible ou nulle sur cette variété ?
2) De façon plus générale, les pathotypes ont-ils une variété de prédilection ?
Les caractères quantitatifs ont été mesurés en serre sur plantes de blé adultes selon la méthodologie décrite par Pariaud et al. (2009 b).
Résultats sur l'évolution des populations...
Évolution rapide
On a analysé 306, 418 et 238 isolats respectivement en 2006, 2007 et 2008.
Sur Soissons et Isengrain, le pathotype 073100 (codé P1), encore prépondérant en 2006 (30-40 %), n'a cessé de décliner par rapport aux années où Soissons dominait le paysage variétal. Il ne représentait plus que 15 % de la population sur cette variété en 2008.
En 2006, une famille composée presqu'exclusivement du pathotype 106314 (codé P4 dans les études de résistance partielle) est devenue dominante. Ce pathotype, apparu en 2003, est surtout inféodé à Caphorn et Trémie-Charger. Mais sur Apache et Orvantis, c'est la famille 006, elle aussi apparue récemment (2002), qui est devenue la plus fréquente.
L'évolution a continué en 2007 : les familles de pathotypes 106314 et 006 sont devenues dominantes quasiment sur toutes les variétés.
La famille de pathotypes 126-136 (pathotypes complexes avec 12 à 13 virulences) qui représentait environ 20 % de la population en 2007, a régressé (6 %) en 2008. En même temps, la famille 106314 a poursuivi sa très forte progression sur toutes les variétés : elle a doublé sa fréquence par rapport à 2007 (aux dépens de la famille 006) pour représenter 70 à 80 % de la population sur Caphorn et Aubusson.
Côté variétés
La modification de la composition de la population pathogène sur la variété Orvantis s'est accompagnée d'une évolution du comportement de cette variété dans le réseau d'essais Arvalis, depuis la résistance en 2001 jusqu'à une sensibilité totale en 2007 (Figure 1).
... sur les composantes de la résistance partielle...
Efficacité de l'infection
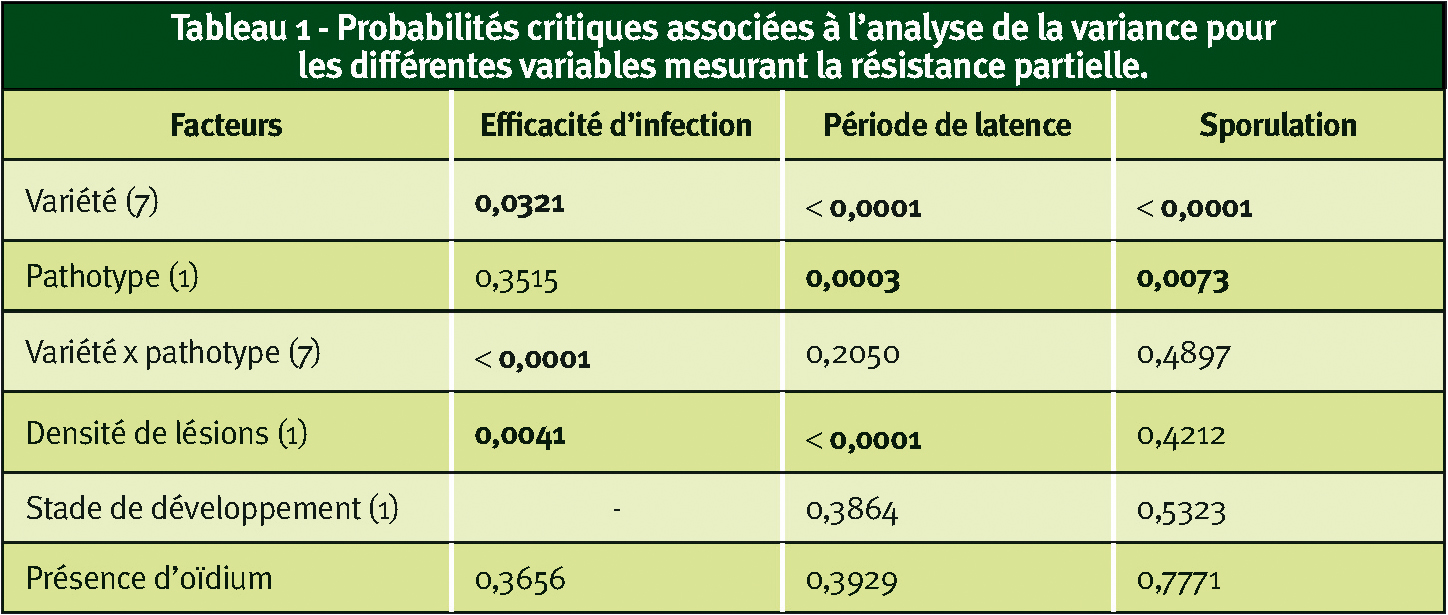
Les résultats de l'étude en conditions contrôlées montrent que la variété, la densité des lésions et l'interaction variété x pathotype ont un effet significatif sur l'efficacité d'infection (ei), mais pas le pathotype (Tableau 1).
L'ei du pathotype P3 est significativement meilleure sur Soissons (0,75) que sur Morocco (0,20, P = 0,0068) et sur Trémie (0,21, P = 0,0005). L'ei du pathotype P4 est significativement supérieure sur PBI (0,62) par rapport à Apache (0,21, P = 0,05). L'interaction variété x pathotype est due au fait qu'une différence significative (P = 0,001) entre P3 et P4 n'existe que pour Trémie, l'ei du P4 (0,37) étant supérieure à celle du P3 (0,21). L'ei du P1 sur Soissons, notre référence pour une interaction compatible très agressive, est équivalente à celle du P3 et du P4. Les ei moyennes, toutes variétés confondues, pour P1, P3 et P4 sont de 0,34, 0,35 et 0,39 respectivement. Sur Morocco, témoin sensible, les ei des deux pathotypes sont faibles (0,20 pour P3 et 0,28 pour P4) et pas significativement différentes des autres variétés, sauf avec Soissons pour P3.
Période de latence
Le pathotype, la variété et la densité de lésions ont un effet significatif sur la période de latence (T50), mais pas l'interaction variété x pathotype (Tableau 1).
Le pathotype P4 a un T50 significativement plus long (218,9 degrés-jours) que P3 (201,7 dj, P=0,0254). Les deux pathotypes ont un T50 plus long que P1 sur Soissons (164,6 dj). Les variétés où le T50 est significativement supérieur par rapport au témoin sensible Morocco (168,9 dj) sont Apache (P = < 0,0001), LD7 (P = < 0,0001) et PBI (P = 0,0014), avec des moyennes de 225, 241 et 207,6 dj respectivement.
Sporulation
La sporulation a été significativement différente entre pathotypes et entre variétés (Tableau 1), mais la densité de lésions et l'interaction variété x pathotype n'ont pas eu d'effet significatif.
Le pathotype P4 a une sporulation plus importante que P3 (P = 0,0086) avec des moyennes de 99,2 ìg/mm² et 92,2 ìg/mm² respectivement. Les deux pathotypes ont une sporulation moins élevée que P1 sur Soissons (122,4 ìg/ mm²). Andalou est la seule variété à sporulation significativement moins forte que Morocco (P = 0,0012) avec des moyennes de 76,1 ìg/ mm² et 104,9 ìg/mm² respectivement.
... et sur l'agressivité de P. triticina et son adaptation à son hôte
Adaptation du pathotype P1 à la variété Soissons
Ce travail est décrit en détail dans la publication de Pariaud et al. (2009 b). Nous avons montré que le pathotype P1 (073100), majoritaire sur Soissons dans les populations naturelles depuis 1999, est plus agressif que les deux autres pathotypes minoritaires P2 (014103) et P3 (166336). Les différences d'agressivité sont de 10 à 25 % environ selon les paramètres d'agressivité considérés (Figure 2).
Étude de l'adaptation différentielle pour les caractères quantitatifs
En utilisant toujours la même méthodologie que ci-dessus, nous avons mesuré l'adaptation quantitative des pathotypes P1, P2 et P3 à diverses variétés. En raison de leurs différences pour les virulences, le test de l'adaptation différentielle de ces pathotypes aux variétés a été fait sur des séries de variétés différentes pour chacun des 3 pathotypes.
L'agressivité du pathotype P1 a été mesurée sur Soissons d'une part, et sur trois autres variétés sensibles sans gènes de résistance spécifique d'autre part (Scipion, Morocco et Festival).
L'agressivité du pathotype P2, supposé adapté à la variété Thésée, a été comparée sur Soissons, Morocco, Thésée et Altria.
Enfin le pathotype P3, originaire de la variété Trémie, a été évalué pour son agressivité sur Soissons et Trémie.
Nous avons montré (figure disponible dans notre communication à la CIMA) que P1 est significativement plus agressif sur sa variété d'origine, Soissons, pour la période de latence, la taille de lésion et la quantité de spores produite.
Les composantes de l'agressivité du pathotype P2 sont toujours dans les plus élevées sur Thésée (taille de lésion significativement plus élevée pour cette variété). Cet agent pathogène a donc bien la capacité à s'adapter à son hôte d'origine pour des caractères quantitatifs.
En revanche cette adaptation ne s'est pas produite pour le pathotype P3 : il se montre au contraire significativement moins performant sur sa variété d'origine Trémie ! Ce résultat est cohérent avec l'observation de la disparition de ce pathotype des populations naturelles seulement 3 ans après son apparition. Ce pathotype complexe doté de 13 virulences s'est comporté comme un « généraliste » capable d'attaquer de nombreuses variétés mais peu agressif, alors que le pathotype P1 adapté à Soissons s'est maintenu fortement dominant sur cette variété, à laquelle il est resté inféodé puisqu'il n'avait pas les virulences nécessaires pour attaquer les variétés plus récentes.
Discussion : efficacité et durabilité des résistances spécifiques...
Le cas d'Orvantis : intérêt du réseau de suivi
L'étude des populations de P. triticina sur 2006-2008 a mis en évidence la forte évolution des populations par rapport aux années 1999-2005, avec la prédominance de deux familles (106314 et 006) qui cumulent les virulences correspondant aux gènes de résistance des principales variétés cultivées (Lr10, Lr13, Lr14a, Lr37).
Ce résultat illustre les capacités d'évolution de l'agent pathogène. On note un cas classique de contournement de gène de résistance spécifique : Orvantis, porteur des gènes Lr10, Lr13 et Lr37 a été protégé par Lr37 jusqu'à l'apparition de pathotypes virulents 37 en 2000.
Le réseau de suivi des populations de P. triticina, fruit de la collaboration de divers partenaires recherche, sélection et développement, est un outil indispensable pour prévoir l'efficacité des résistances de type spécifique : il renseigne sur les combinaisons de virulences présentes dans la population ainsi que sur leur fréquence.
Il reste cependant difficile de prédire la durabilité de combinaisons de gènes de résistance spécifique. En effet, des combinaisons de virulence peuvent apparaître mais ne pas augmenter en fréquence (voire disparaître) en raison d'une faible agressivité.
Le cas de Caphorn : intérêt de mesurer l'agressivité des pathotypes
Ceci peut être illustré par le cas de Caphorn : cette variété est sensible aux pathotypes de la famille 126-136, mais nous avons montré que la fréquence de cette famille se maintient au champ à un niveau relativement faible sans doute en raison d'une agressivité moindre.
Ceci pourrait expliquer le comportement de Caphorn dans le réseau Arvalis : la variété semble évoluer vers une moindre résistance. Elle exprimerait un niveau de maladie élevé dans quelques essais où cette famille de pathotypes est présente mais resterait bien résistante dans la majorité des essais en raison de la faible fréquence de la famille 126-136.
À partir de 2008...
Nous avons voulu compléter l'information apportée par les suivis de population à l'aide d'essais variétaux mis en place par Arvalis à partir de 2008, chaque essai étant inoculé par un seul pathotype. L'objectif premier de ces essais est d'évaluer la résistance partielle des variétés, mais ils permettent aussi de confirmer (ou non) au champ les informations acquises en conditions contrôlées sur la résistance spécifique.
Les combinaisons de gènes de résistance spécifique restent une ressource à exploiter et leur efficacité peut être estimée grâce au réseau de suivi des populations. Mais leur durabilité n'est pas toujours facile à évaluer car on manque de données sur l'agressivité des pathotypes.
... efficacité et durabilité des résistances partielles
Analysable au laboratoire
La méthodologie utilisée a permis de retrouver en conditions contrôlées la résistance partielle préalablement détectée au champ, et d'en analyser les composantes. Nous avons ainsi mis en évidence que la résistance partielle des différentes variétés et lignées étudiées s'exprimait sur différentes composantes : elle se traduit par une réduction de l'efficacité d'infection du pathotype P3 sur Trémie, une augmentation de la période de latence de P3 et P4 sur Apache, LD7 et PBI, et une réduction de la production de spores de P3 et P4 sur Andalou.
Ces résultats montrent que l'on peut évaluer la résistance partielle en conditions contrôlées... mais les mesures restent assez lourdes.
Compter avec les pathotypes. Et les années
De plus, l'existence d'une interaction variété x pathotype pour certaines composantes suggère que des relations de type spécifique existent aussi pour les résistances de type quantitatif. Le niveau de résistance partiel d'une variété peut donc varier selon le pathotype auquel elle est confrontée.
En l'absence de connaissances plus précises sur le déterminisme génétique des sources de résistance présentes dans le matériel génétique utilisé en France, ces mesures permettent d'assurer une diversification des sources en recherchant des résistances basées sur des composantes diversifiées : efficacité d'infection, latence, sporulation...
Nous avons également montré que les pathotypes présentent un certain niveau de spécialisation pour un fonds génétique de l'hôte donné : il existe donc une adaptation différentielle pour les caractères quantitatifs. Il s'ensuit un risque d'érosion de la résistance quantitative en cas de culture à grande échelle. D'où l'importance de conserver un objectif de diversification des sources de résistances pour préserver la durabilité des résistances de type quantitatif.
Conclusion
L'utilisation de toutes les ressources génétiques de résistance à la rouille brune disponibles – nécessaire pour limiter le risque épidémique sur blé – doit s'accompagner d'une vigilance concernant la résistance de type spécifique. Ceci exige la pérennité du réseau de suivi des virulences dans les populations associant recherche, sélection et développement pour un maximum d'efficacité. Il sera utilement complété par des essais variétaux (Arvalis-INRA) avec pathotypes séparés pour améliorer l'estimation de la durabilité.
<p>* INRA UMR BIOGER - BP 01 - 78850 Thiverva-Grignon goyeau@grignon.inra.fr</p> <p>** Arvalis-Institut du Végétal, Chemin de la Petite-Minière, 78280 Guyancourt. *** Arvalis-Institut du Végétal, 91720 Boigneville.</p> <p>(1) En 2007, on a pu évaluer à 23 q/ha en moyenne la perte de rendement en l'absence de traitements (essais variétés d'Arvalis-Institut du végétal de cette année-là).</p> <p>(2) Centre international pour l'amélioration du maïs et du blé.</p> <p>(3) Quantitative Trait Locus = lieu (du chromosome) porteur de caractère (trait) quantitatif.</p> <p>(4) Par agressivité, on entend l'importance quantitative des symptômes produits sur une plante sensible par un individu parasite. Dans le cas de la rouille brune, parasite biotrophe strict, la notion d'agressivité se confond avec celle de fitness, telle qu'elle est utilisée en génétique des populations (Pariaud et al., 2009 a).</p> <p>Une méthodologie d'étude des composantes de la résistance partielle en serre sur plantes adultes est désormais au point. Mais elle reste assez lourde et ne peut être appliquée qu'à un nombre limité de combinaisons pathotype x variété.</p> <p>Cette même méthodologie a permis de montrer l'existence d'une capacité d'adaptation chez <i>P. triticina</i> à la résistance quantitative. Ceci suggère que la résistance partielle, réputée davantage durable que la résistance spécifique, n'est pas absolument et éternellement durable en soi... Il faut préserver sa durabilité en diversifiant les sources de résistance utilisées et en les associant à des résistances de type spécifique.</p> <p><b>Remerciements :</b> Le financement de cette étude a été en grande partie assuré par le CAS DAR 391 <i>« Mise au point et utilisation d'outils de biovigilance et de gestion durable des résistances variétales aux pathogènes ».</i> Les auteurs tiennent également à remercier le GNIS-FSOV (Fonds de soutien à l'obtention végétale), qui a également fortement contribué au financement des travaux présentés. Enfin et surtout, un grand merci à tous les collaborateurs du réseau de collecte des échantillons de rouille pour leur collaboration efficace et... durable.</p>
Figure 1 -
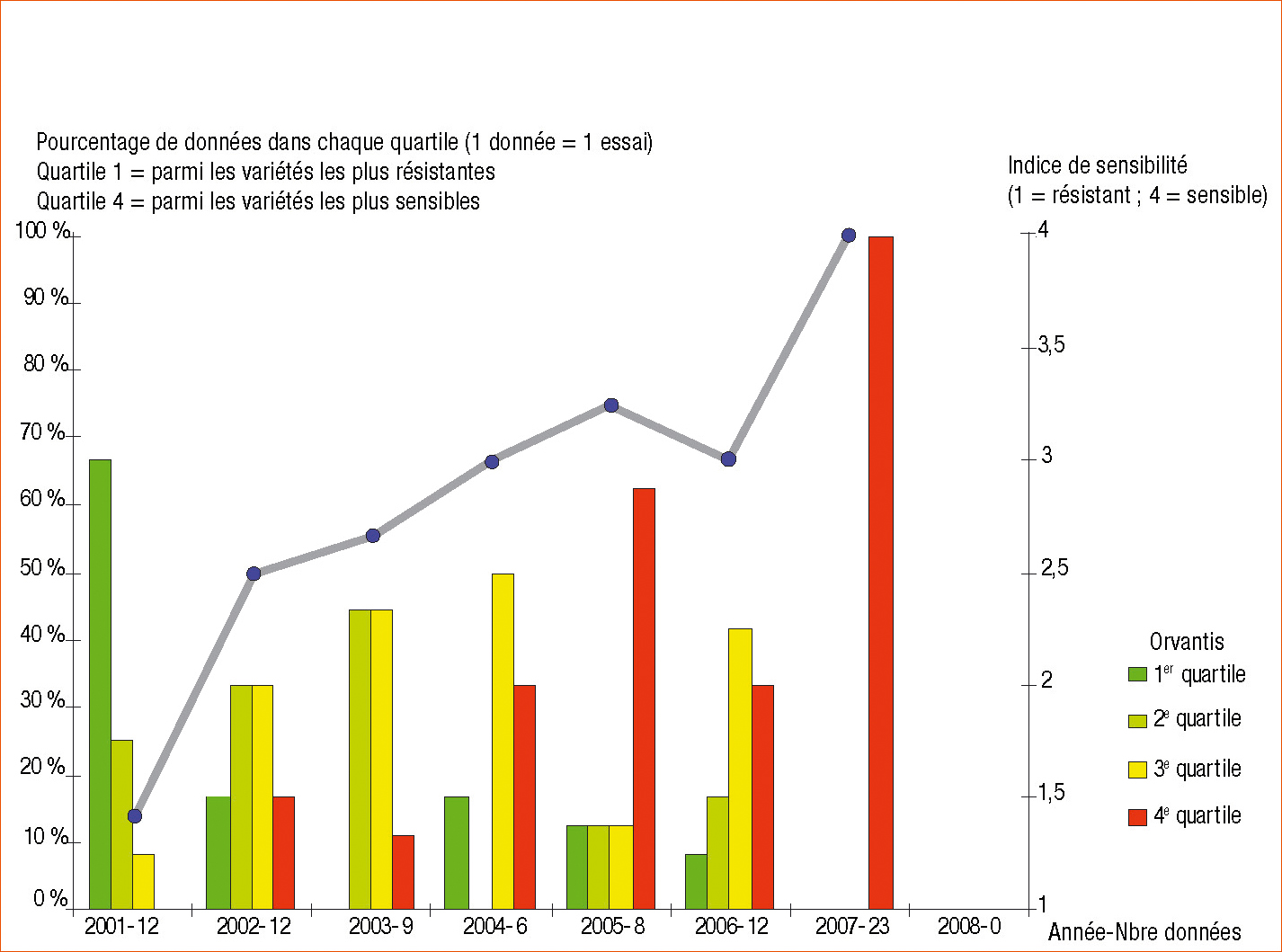
Évolution de la sensibilité d'Orvantis dans le réseau d'essais Arvalis sur la période 2001-2008.
Figure 2 -


Symptômes après inoculation artificielle en serre sur plantes adultes. ph. B. Pariaud
Mesure de l'agressivité relative des pathotypes de P. triticina P1, P2 et P3 sur la variété Soissons, pour différentes variables.

