
 Imprimer
Imprimer
ph. B. Chaubet, INRA
, peut aussi toucher le blé, comme ci-contre. Le modèle présenté ici pourrait permettre de prévoir les risques en la matière. Si des pièges peuvent subsister pour le « nourrir » ! ph. F. Fabre, INRA")
On parle toujours de la JNO comme de la jaunisse nanisante de l'orge mais son agent, le BYDV (Barley Yellow Dwarf Virus), peut aussi toucher le blé, comme ci-contre. Le modèle présenté ici pourrait permettre de prévoir les risques en la matière. Si des pièges peuvent subsister pour le « nourrir » ! ph. F. Fabre, INRA
Pour raisonner la protection de ses cultures, l'agriculteur peut utiliser des OAD, outils d'aide à la décision. Parmi eux, certains modèles permettent de prévoir des risques phytosanitaires à partir de données climatiques recueillies automatiquement. Mais d'autres, pour « tourner », utilisent des données telles que l'arrivée d'un ravageur ; il en est de même pour la simple règle de décision « traitement à partir d'un seuil d'infestation donné ». Mais qui peut fournir ces informations ? Libérer l'agriculteur ou son conseiller des observations au champ, est-ce possible ? Nous proposons ici une solution, c'est... un modèle ! Il utilise les données du réseau de piégeage Agraphid. Qui est à sauver d'urgence.
L'accroissement des contraintes économiques et environnementales pesant sur la production agricole incite les agriculteurs à raisonner au mieux les interventions phytosanitaires. Dans ce contexte, les outils d'aide à la décision (OAD) procurent une aide précieuse. En théorie tout au moins...
Libérer l'agriculteur des observations au champ
Car en pratique, de nombreux OAD doivent être initialisés avec des données de terrain acquises à l'échelle de l'exploitation, voire à celle de la parcelle. Cette étape préalable de collecte de l'information rebute souvent les agriculteurs qui n'en ont ni le temps ni parfois la formation. Ainsi, bien des OAD tombent dans les oubliettes d'un coin de disque dur. Et ils s'y laisseront oublier tant que n'auront pas vu le jour des solutions remplaçant tout ou partie des notations de terrain par des moyens indirects d'observation.
Dans le cas des insectes ravageurs, en particulier des pucerons, plusieurs pays européens dont la France disposent ou disposaient d'un réseau de pièges à aspiration qui échantillonnent quotidiennement selon un protocole standardisé les populations d'insectes en déplacement aérien (Encadré 1).
Ce qui manquait au réseau de pièges pour les « avertissements »
Outre leur intérêt scientifique pour l'étude de migrations d'insectes, d'invasions biologiques et de l'évolution de la biodiversité (liée au changement climatique, etc.), ces pièges avaient aussi été conçus comme des outils d'avertissements agricoles (Robert & Choppin de Janvry, 1977).
Mais pour ce faire une phase préalable d'étalonnage est nécessaire. Elle consiste à établir une relation statistique robuste entre, d'un côté, la quantité de pucerons piégés et, de l'autre, celle infestant les cultures situées dans un rayon allant jusqu'à une centaine de kilomètres du piège. Or, malgré des tentatives préliminaires (Gamon, 1989 ; Dedryver et al., 1991), aucun travail de recherche n'avait établi solidement de relation de ce type.
Ce manque est dû en grande part au fait que l'opération exige de confronter d'importants jeux de données de piégeage (ils existent) avec des données issues de comptages sur le terrain. Ces derniers sont d'autant plus rares qu'ils doivent être pluriannuels et plurirégionaux afin de représenter la variabilité des processus étudiés. De ce fait, on était encore il y a peu incapable de modéliser la contamination d'un champ « moyen » à partir des données fournies par le piège à aspiration le plus proche. Ce fut la cause probable du désintérêt de nombreux acteurs du développement agricole pour le réseau français de ces pièges cette dernière décennie.
Un récent travail comble ce manque
Un récent travail effectué sur Rhopalosiphum padi, puceron des céréales vecteur principal de la Jaunisse nanisante de l'orge (JNO), nous a permis de démontrer pour la première fois que les données de capture de pucerons aux pièges à aspiration procurent, par l'intermédiaire d'un modèle ad hoc, de bonnes estimations des taux de contamination des champs par les pucerons ailés en automne quelles que soient la région et l'année. Ce modèle constitue une alternative pratique et fiable aux comptages fastidieux (souvent à quatre pattes !) et dispendieux des insectes ravageurs dans les champs.
Présentation du modèle
Un peu de technique...
Dans cette étude (Fabre et al., 2010), nous avons utilisé deux jeux de données. Le premier est constitué par 387 données de comptage hebdomadaire de R. padi ailés entre fin septembre et fin novembre, recueillies sur 66 parcelles d'orge d'hiver implantées par la société Bayer CSF dans la moitié nord de la France entre 1989 et 2002. Chaque comptage a été réalisé sur 400 plantes par parcelle.
Le second jeu de données est constitué de captures journalières de R. padi en automne par le réseau de pièges à aspiration. À chaque comptage parcellaire a été associé le nombre de R. padi ailés capturés au piège à aspiration le plus proche de la parcelle durant les 3 semaines précédant la date d'échantillonnage.
Puis un modèle bayésien hiérarchique (HBM) décrivant la colonisation des céréales d'hiver par les pucerons ailés de l'espèce R. padi a été ajusté à ces données. Il permet d'inférer, par l'introduction de variables « latentes », les valeurs de processus et de paramètres du modèle non mesurables directement (encadré 2 page suivante), comme la densité de pucerons volant au-dessus d'un champ ou atterrissant dans celui-ci ou encore la probabilité qu'ils survivent une fois posés.
... et trois résultats notables
Les principaux résultats à retenir sont : 1) La valeur prédictive du modèle, estimée à l'aide de tests de validation croisée, est bonne : le coefficient de détermination (R2) entre les nombres de R. padi ailés observés dans les champs et leurs valeurs prédites par le modèle à partir des captures aux pièges à aspiration le plus proche est de 0,53 (Figure 1).
Par ailleurs 97 % des valeurs prédites se trouvent dans un intervalle de confiance à 95 % ; il s'agit d'un autre avantage majeur de l'usage des modèles bayésiens pour l'aide à la décision : permettre une quantification explicite de l'incertitude associée à des prédictions.
2) Le taux de déposition, c'est-à-dire la relation entre le nombre de pucerons volant au-dessus du piège à aspiration et la quantité colonisant effectivement les champs, est corrélé négativement avec la sévérité locale de l'hiver mesurée par le nombre moyen de jours où la température minimale est inférieure à -5 °C (Figure 2).
Nous vérifions qu'à quantité égale de pucerons en migration, la colonisation d'une parcelle (donc le risque JNO) diffère selon sa localisation géographique : en effet la proportion de clones sexués dont les ailés volent en automne sans coloniser les jeunes céréales (ils migrent vers les merisiers à grappes) donc sans participer à l'épidémiologie de la JNO, est d'autant plus forte que les hivers sont rigoureux (Encadré 3).
3) Selon le modèle, la probabilité de présence d'un puceron ailé dans un champ décroît fortement après son atterrissage et moins de 5 % des immigrants restent présents dans la culture 10 jours après leur arrivée. A priori, ces disparitions résultent surtout de la mortalité des pucerons et, marginalement, de leur émigration.
Applications pratiques : espoirs et contraintes
Nourrir un OAD « anti-JNO »
L'application directe de cette étude concerne l'OAD « aphi.net® » de gestion des traitements contre les pucerons vecteurs de la JNO développé par l'INRA et distribué par Bayer CSF (Dedryver et al., 2010). Cet OAD requiert un comptage de pucerons au champ pour initialiser un modèle calculant les probabilités de dégâts en fonction de la température de l'automne. Ce comptage pourrait être avantageusement remplacé par une prévision d'infestation calculée en temps réel à partir des données régionales de piégeage. Pour l'heure le couplage des modèles et leurs implémentations dans le logiciel existant n'est toutefois pas encore effectif.
D'autres espèces ? Pas toutes
L'étape suivante consisterait à transposer cette étude à d'autres espèces de pucerons, voire à d'autres insectes comme les cécidomyies des fleurs de blé (Bayon et al., 1983). Le défi reste à relever.
En effet, le modèle relie deux séries de données issues, l'une de nombres d'insectes capturés par les pièges à aspiration et l'autre de comptages au champ. Les espèces piégées dans le réseau Agraphid et leur représentativité géographique ne sont pas limitantes car l'essentiel du réseau a fonctionné jusqu'en 2002. En revanche, il faut des jeux de données de terrain recueillies entre 1980 et 2002. Or seuls l'industrie phytosanitaire et quelques instituts techniques en disposent, surtout sur grandes cultures (céréales, maïs, colza, voire betteraves).
Par ailleurs, le modèle ne fonctionne sous sa forme actuelle que si le vol de colonisation des parcelles est limité dans le temps et dissocié du vol d'émigration (il ne faut pas confondre, parmi les insectes piégés, ceux qui « décollent » et ceux qui atterrissent !).
Enfin les captures de l'espèce choisie doivent être assez abondantes pour se prêter à un traitement statistique.
Sur la base de ces critères, il est envisageable de modéliser la contamination des maïs en juin-juillet par les 3 principales espèces de pucerons issues du blé et de l'orge (Sitobion avenae, R. padi et Metopolophium dirhodum).
Concernant le puceron « des épis » sur blé au printemps (S. avenae), il existe un continuum entre l'immigration (arrivée d'ailés dans la parcelle) et l'émigration (envol d'ailés produits dans cette parcelle). Toutefois cette difficulté est surmontable car nous avons mis en évidence une corrélation entre les nombres de S. avenae piégés et ceux atterrissant sur une culture de blé pendant une période de 20 à 30 jours autour de l'épiaison (Vialatte et al., 2007). Ces résultats valables localement (région de Rennes) exigent d'être confirmés à plus grande échelle.
Trois autres espèces de pucerons respectent également ces critères : Myzus persicae sur betteraves, Acyrthosiphon pisum sur pois de printemps et Brevicoryne brassicae, surtout en cultures de choux de printemps ou d'été.
Mais il faut garder les pièges !
Par ailleurs, même s'il s'apparente à une lapalissade, il y a un écueil à l'application concrète de ces recherches à ne pas sous-estimer.
En effet, ces modèles peuvent être construits avec des jeux de données historiques mais ils ne fonctionneront qu'avec des données de piégeage actuelles, fournies jour après jour par un réseau de pièges à aspiration fonctionnel !
Or la survie d'Agraphid est hélas loin d'être l'hypothèse la plus probable actuellement : si 5 pièges existent toujours, seuls 2 (Le Rheu, Poitiers) fonctionnent en permanence tandis que 3 (Auxerre, Orléans et Reims) ne tournent « qu'à la demande », surtout des firmes phytosanitaires. Un 6e piège est en suspens à Avignon.
Un effort doit être fait rapidement pour pérenniser ce dispositif et, si possible, le compléter par un ou deux pièges supplémentaires (dans le sud-ouest et en Bourgogne ou Lorraine) afin d'assurer une couverture satisfaisante de nos principales zones de grandes cultures.
Pour finir, si le « confort d'utilisation » est un volet important du cahier des charges d'un OAD, sa qualité prédictive, mesurée par sa spécificité et sa sensibilité, en est un autre car elle conditionne l'intérêt économique qu'en tirent les agriculteurs (Fabre et al., 2007). C'est donc aussi sur cette composante des OAD qu'il faut agir, en continuant les recherches en amont, et, en ce qui nous concerne, en élargissant l'exploitation des données de piégeage d'insectes à la simulation des phases ultérieures de leurs dynamiques de population.
<p>* INRA/Agrocampus-Ouest/Université Rennes1, UMR BiO3P : charles-antoine.dedryver@rennes.inra.fr, maurice.hulle@rennes.inra.fr, manuel.plantegenest@agrocampus-ouest.fr</p> <p>** INRA/Agrocampus-Ouest, UMR Écologie et Santé des écosystèmes. etienne.rivot@agrocampus-ouest.fr</p> <p>*** INRA, UR 407, Unité Pathologie végétale. frederic.fabre@avignon.inra.fr</p>
1 - Les réseaux de piégeages à aspiration

Piège à aspiration. J. Bonhomme/INRA

Le réseau EXAMINE en juin 2009 http://www.rothamsted.bbsrc.ac.uk/examine/
.")
Les pièges du réseau Agraphid en fonction ou ayant fonctionné (en rouge ceux dont les captures ont été utilisées pour la présente étude ; les deux gros points représentent les pièges fonctionnant encore en 2010).
Les pièges à aspiration (photo) fournissent des données quotidiennes de l'activité de vol des pucerons devant permettre d'anticiper la colonisation des champs. Cette information est particulièrement utile dans l'évaluation des risques liés aux maladies à virus.
Fluctuations médiatisées
Ces données à vocation régionale permettent aussi d'étudier les fluctuations des populations de pucerons depuis plus de 40 ans en Europe et d'évaluer l'impact des changements globaux sur leur biodiversité et leurs rythmes d'activité.
On a ainsi observé une augmentation du nombre d'espèces présentes en France en raison d'une part d'introductions fréquentes liées à l'augmentation des échanges internationaux de matériel végétal et, d'autre part, de l'activité accrue d'espèces qui étaient déjà présentes sur notre territoire mais à des niveaux auparavant trop faibles pour être détectés. De même, une augmentation de la précocité printanière des vols a été mise en évidence. L'avance est d'un jour par an en moyenne depuis 30 ans.
Ces résultats ont été publiés (Hullé et al., 2010) et médiatisés (presse grand public).
Agraphid(1) est la composante française du réseau EXAMINE. Ce réseau mis en place en 1978 à partir de 5 pièges s'est étoffé au fil des années pour atteindre 15 pièges. Actuellement il reste 2 pièges en fonctionnement permanent.
(1) Créée en 1978 par l'Institut national de la recherche agronomique (INRA), l'Association de coordination technique agricole (ACTA) et les Services régionaux de la protection des végétaux (SRPV).
Figure 1 - Valeur prédictive du modèle estimée à l'aide de tests de validation croisée.
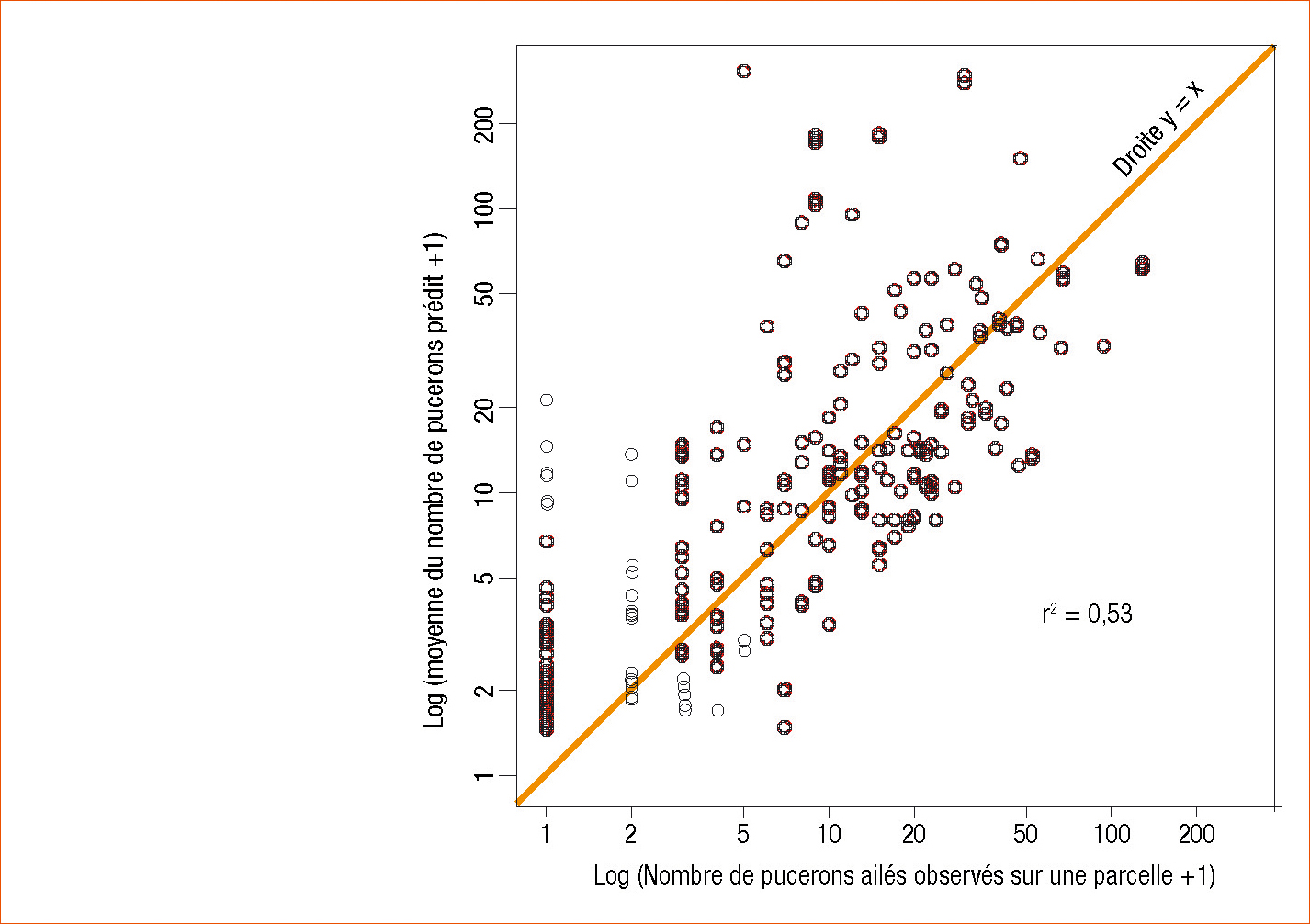
Le graphe représente la corrélation entre des valeurs observées de nombre de pucerons ailés sur une parcelle (en abscisse) et des valeurs prédites par le modèle de ce même nombre (en ordonnée).
Le coefficient de détermination (R²) entre les deux est de 0,53. La validation croisée est une méthode d'estimation de la fiabilité d'un modèle qui permet de dissocier données de « construction » et données de « validation ». En effet, les performances futures d'un modèle ne peuvent pas être estimées correctement sur les données qui ont servi à le construire.
2 - De l'intérêt des modèles bayésiens hiérarchiques en écologie
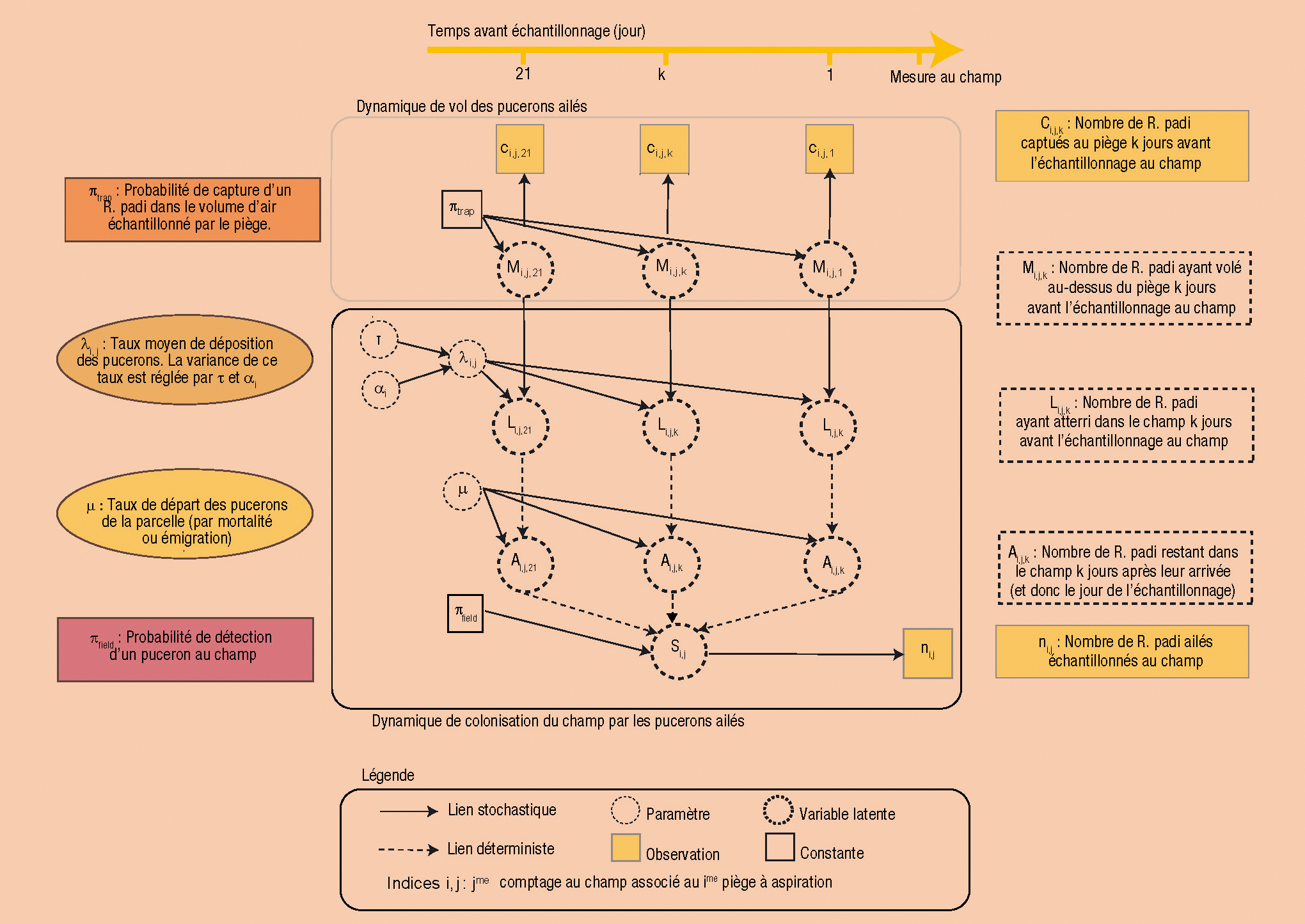
Au cours des 20 dernières années, les modèles bayésiens hiérarchiques ont connu un bel essor dans le domaine de l'écologie statistique. En effet, ils permettent de lever certains verrous méthodologiques des approches plus classiques de modélisation quantitative.
La modélisation hiérarchique est fondée sur la description du système à modéliser en différentes « couches ». Celles-ci correspondent aux paramètres du modèle, aux variables d'état latentes du système (variables pertinentes pour décrire le système mais non observables) et aux observations traduisant, elles, des manifestations indirectes des processus latents d'intérêts.
La modélisation hiérarchique relie ces couches entre elles par une structure de conditionnement probabiliste qui capture les relations de cause à effet entre les variables. En plus d'être conceptuellement satisfaisante, cette structure générale apporte de la flexibilité car elle distingue explicitement les processus écologiques des processus d'observation et sépare les sources de variabilité et d'incertitude associées à chaque « couche ».
Ainsi il est possible de représenter la complexité des processus écologiques comme des emboîtements d'échelles (individus, populations, ensembles de populations), des relations induites par les dimensions dynamiques et/ou spatiales ainsi que les différentes sources de variabilité et de d'incertitude associées.
Par ailleurs, la connexion avec les modèles d'observation facilite l'intégration de sources d'information multiples issues de méthodes d'échantillonnage et de suivi des populations naturelles.
Le modèle présenté dans cet article est un exemple de modèle bayésien hiérarchique. Le diagramme ci-dessous permet de distinguer clairement l'articulation entre les couches du modèle (cf légende).
Ainsi, il est impossible d'observer directement au champ l'arrivée de pucerons ailés. En revanche on peut réaliser un comptage au champ du nombre de pucerons présents à un instant donné. Ce comptage (variables observée ni,j, dans le diagramme) est, à une erreur d'observation près, πfield, la manifestation indirecte de la colonisation de la parcelle par les pucerons durant les journées ayant précédé ce comptage en prenant en compte que, plus on remonte loin dans le temps, plus il est probable que les pucerons aient disparu le jour de l'observation (par mortalité ou émigration) (paramètre ì - variables latentes Ai,j,k et Si,j).
L'intensité quotidienne de cette colonisation dépend à son tour de l'intensité des vols de pucerons (variables observée ci,j,k mesurée à une erreur près, πtrap, par les pièges à aspiration et variables latentes Mi,j,k et Li,j,k) et d'un taux de déposition qui dépend de la localisation du piège (paramètres αi).
Figure 2 - Relation entre la sévérité de l'hiver (ici le nombre moyen de jours où la température minimale est inférieure ou égale à – 5 °C) et le « taux de déposition des pucerons » sur les semis d'orge.
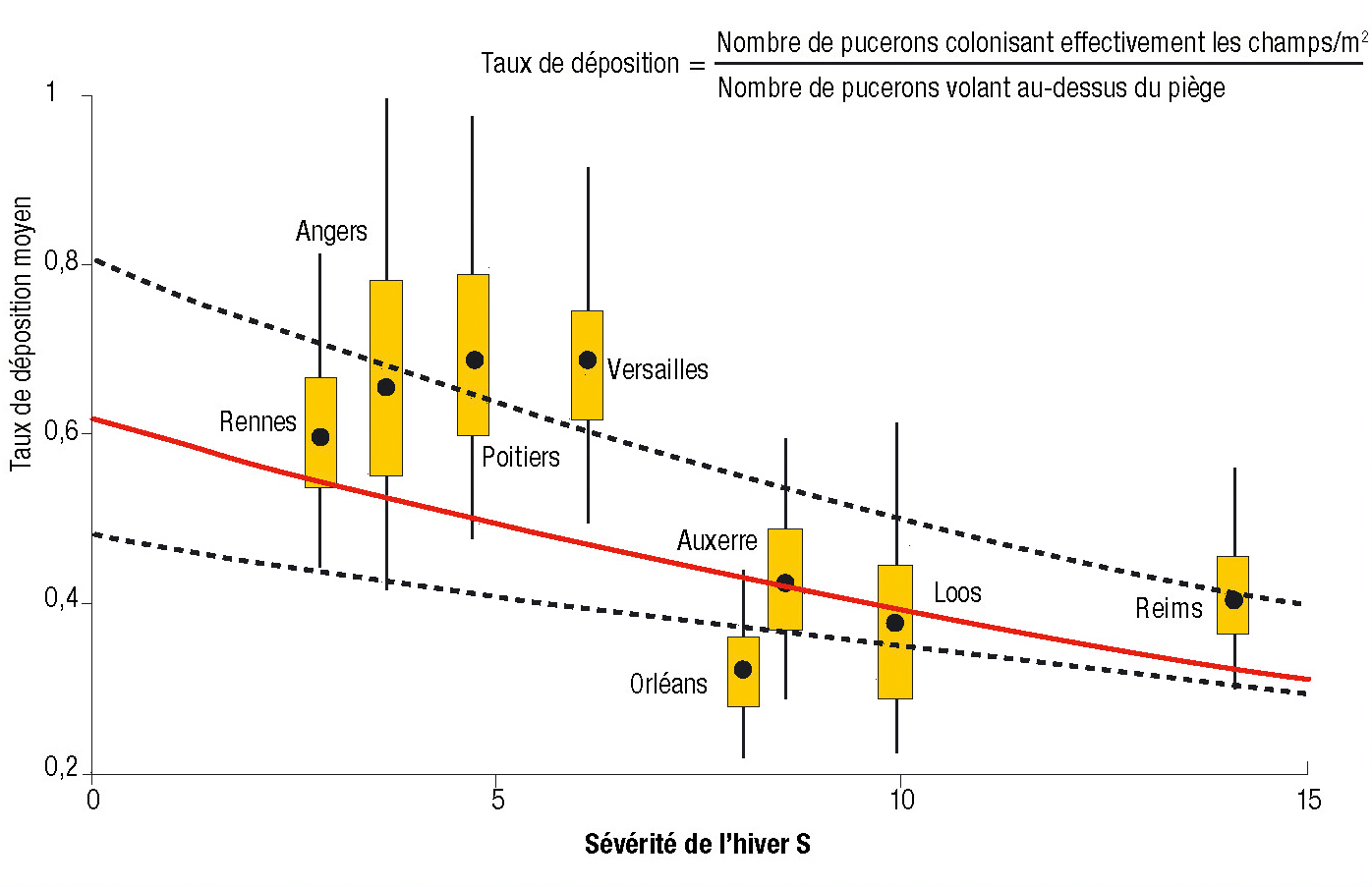
Pour chaque piège à aspiration, une boîte à moustache indique la valeur moyenne du taux de déposition (•), les quantiles 25 % et 75 % (rectangle) ainsi que les quantiles 5 et 95 % (extrémités des segments).
Un test de corrélation de Pearson détecte une corrélation négative entre la sévérité de l'hiver et les taux de déposition des pucerons (t=-0.257, d.f.=6, P=0.02). Par ailleurs, une seconde approche non détaillée ici (cf Fabre et al., 2010) permet de modéliser directement le lien entre ces deux variables. La ligne continue décrit cette relation, les lignes pointillées indiquant elles la bande de confiance à 95 %.
3 - La biologie du puceron Rhopalosiphum padi

R. padi ailé. S'il s'agit d'une femelle parthénogénétique, elle est capable de migrer sur les céréales à l'automne. ph. B. Chaubet, INRA
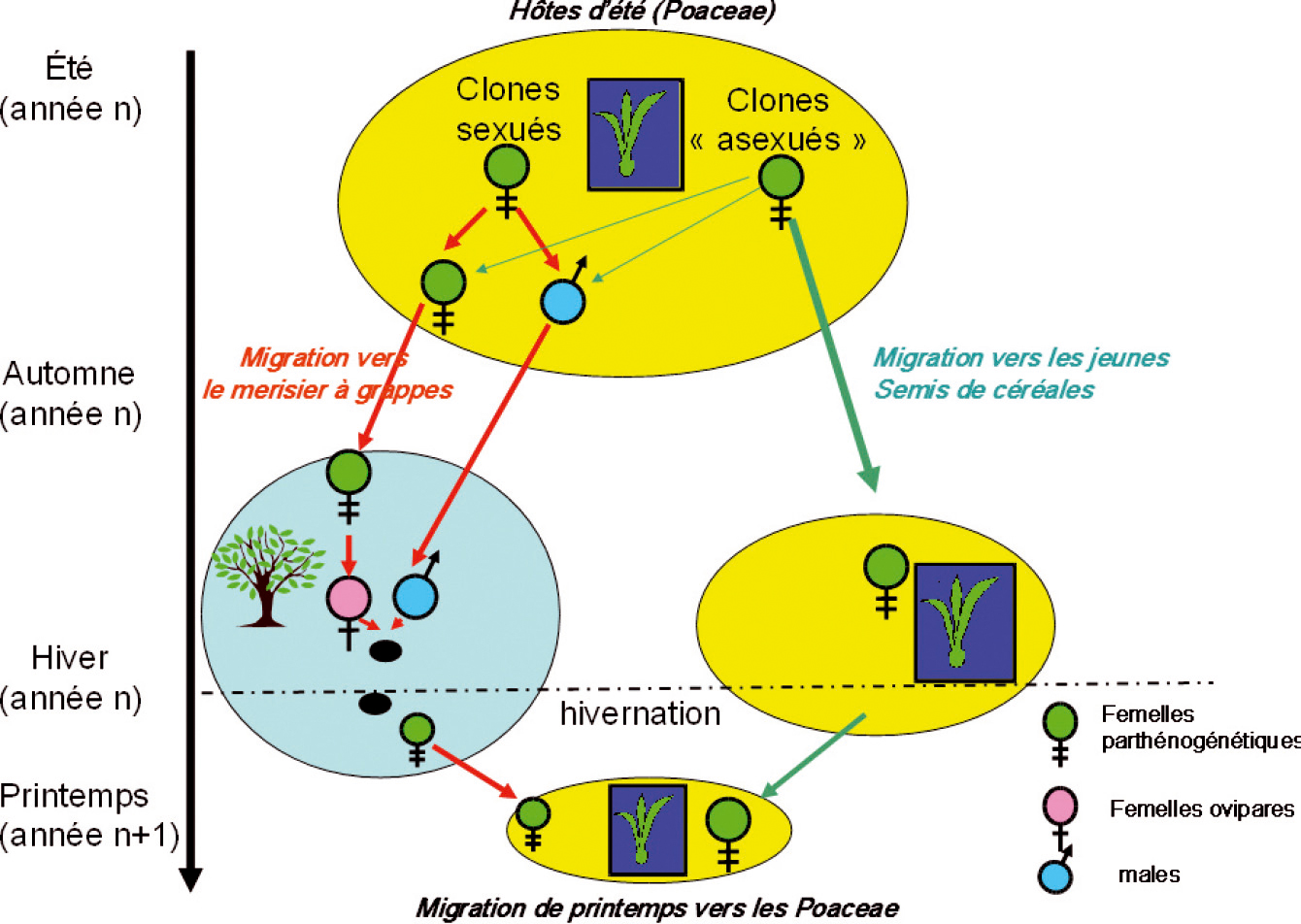
R. padi, en cours d'année, alterne entre les graminées (Poacées) où il développe de nombreuses générations parthénogénétiques pendant la belle saison, et un arbuste, le merisier à grappes (Prunus padus), où les femelles de l'unique génération sexuée annuelle pondent des œufs fécondés qui diapausent durant l'hiver et éclosent au printemps suivant. Les femelles parthénogénétiques qui en sont issues développent des colonies sur l'arbuste. Puis ces colonies donnent naissance courant mai à des ailés migrant vers les graminées.
Néanmoins, une grande partie des populations de R. padi en France passent outre cette étape de reproduction sexuée sur P. padus en se maintenant tout l'hiver à l'état parthénogénétique sur les graminées.
La figure ci-dessous illustre les deux stratégies de reproduction de R. padi en automne : les lignées sexuées produisent des mâles et des pré-femelles (gynopares) qui migrent obligatoirement vers les merisiers à grappes, alors que les lignées essentiellement asexuées produisent quelques formes sexuées et surtout des femelles parthénogénétiques migrant vers les jeunes céréales. Ainsi les R. padi capturés par les pièges à aspiration en automne sont un mélange de males et pré-femelles migrant vers les merisiers à grappes et de femelles parthénogénétiques migrant vers les graminées disponibles, jeunes céréales essentiellement.
Le rapport entre ces deux catégories varie en fonction des régions car le degré d'investissement de R. padi dans la reproduction sexuée dépend du climat hivernal (les populations parthénogénétiques sont tuées par le froid). Ceci est un paramètre important à prendre en compte car seuls les pucerons parthénogénétiques migrant à l'automne vers les graminées ont un rôle dans l'épidémiologie de la JNO.

