
 Imprimer
Imprimer
Symptômes de mildiou sur feuilles. Trois variétés résistantes issues des travaux de l'Inra et des obtenteurs privés ont été inscrites au catalogue français entre 2008 et 2014. Pour ces trois variétés, le travail avait commencé en 1982. Concernant la résistance au nématode Globodera pallida, ce fut plus rapide : le travail débuta en 1988, la première inscription fut en 2009.. Photo : Euroblight
Plus de quarante bioagresseurs d'importance économique peuvent s'attaquer à la pomme de terre.
Parmi ces parasites et ravageurs (bactéries, champignons, oomycètes, virus, viroïdes, nématodes, insectes, etc.), le mildiou et les nématodes sont particulièrement nuisibles. Un des moyens de les combattre est la lutte génétique.
Pourquoi la lutte génétique ?
Deux cents « cousins » sauvages, dont certains résistants
En effet, la pomme de terre (Solanum tuberosum) fait partie de la famille des solanacées, originaire d'Amérique du Sud, qui comprend aussi d'autres espèces d'intérêt agronomique (tomate, tabac, aubergine, poivron, etc.), mais également plus de deux cents espèces sauvages produisant des tubercules. Or ces dernières ont coévolué avec les principaux parasites que l'on connaît aujourd'hui sur la pomme de terre cultivée, et différents mécanismes d'évitement ou de résistance leur ont permis de survivre dans la nature face à leurs ennemis.
L'utilisation de ces mécanismes a été très vite envisagée pour améliorer la résistance de la pomme de terre cultivée à ses bioagresseurs.
Culture sensible donc traitée
En effet, la grande sensibilité de la culture à divers parasites et ravageurs impose le recours à de nombreux traitements fongicides, insecticides et/ou nématicides durant le cycle cultural ou lors de la préparation des sols.
La pomme de terre est, avec la vigne et l'arboriculture, une des cultures les plus consommatrices de pesticides à l'hectare avec un indice de fréquence des traitements (IFT) de l'ordre de 16, hors herbicides.
La résistance ou tolérance variétale pour diminuer les traitements
Dans le cadre du plan Ecophyto, il existe depuis 2008 des pressions croissantes sur les acteurs de la filière pour tenter de limiter le recours aux produits phytosanitaires.
Les efforts de recherche des scientifiques et techniciens de l'UMR Igepp (Institut de génétique, environnement et protection des plantes), du centre Inra de Bretagne-Basse-Normandie, s'inscrivent dans ce contexte. Ils ont notamment permis d'identifier des sources de résistance chez des espèces apparentées, d'en étudier les déterminants génétiques et les possibilités de transfert par croisement à l'espèce cultivée.
Nous indiquerons ici les principaux résultats des travaux de génétique et comment ils débouchent aujourd'hui sur la création de matériel végétal innovant. Les nouveaux développements espérés à l'avenir dans ce domaine seront présentés.
Nous nous focaliserons sur les parasites sur lesquels l'Igepp met aujourd'hui ses forces de recherche : le mildiou de la pomme de terre (Encadré 1) et les nématodes (Encadré 2 p. 26).
Des ressources génétiques aux nouvelles variétés : un processus long et complexe
À l'Inra, le screening des collections
Les ressources génétiques sont à la base de l'acquisition de connaissances et du matériel végétal amélioré.
En matière de solanacées tubéreuses, les chercheurs et obtenteurs français disposent d'un matériel abondant et précieux conservé dans le Centre de ressources biologiques (CRB) BrACySol, situé sur le site Inra de Ploudaniel (Finistère).
Les collections y sont caractérisées par de nombreux tests, ce qui a permis d'identifier différentes sources de résistance vis-à-vis des parasites étudiés (Encadré 3 p. 27).
Obtention de « géniteurs améliorés »
Des croisements ont été entrepris pour tenter le transfert par voie sexuée de ces nouvelles résistances à l'espèce cultivée. Dans certains cas, les croisements sont assez faciles à réaliser mais, la plupart du temps, des barrières (niveau de ploïdie, incompatibilité pollinique, stérilité mâle) constituent des freins importants à la réalisation des croisements entre solanacées et doivent être surmontées pour obtenir des hybrides interspécifiques.
Quand ces difficultés ont pu être maîtrisées, il s'ensuit un processus souvent très long de rétrocroisements et de sélection. Il permet d'éliminer progressivement les caractères indésirables du parent sauvage tout en relevant le niveau agronomique et en conservant dans la descendance la ou les résistances ciblées. Au terme de ce processus qui, suivant l'espèce de départ, prend entre dix et vingt ans, la sélection conduit à des génotypes dont les caractéristiques majeures sont celles de la pomme de terre cultivée et qui sont porteurs de gènes de résistance nouveaux. Ces génotypes ne sont pas encore des variétés mais des « géniteurs améliorés » (Figure 1 p. 26).
Quatre obtenteurs français se dirigent vers les variétés commerciales
Dans le cadre d'une convention qui lie l'Inra et l'ACVNPT (Association des créateurs de variétés nouvelles de pomme de terre) depuis plus de quarante ans, les géniteurs sont proposés sous exclusivité aux quatre obtenteurs français qui peuvent les introduire dans leurs programmes de croisements pour créer de nouvelles variétés. Au moins un cycle de croisement supplémentaire et dix ans de sélection sont ensuite nécessaires pour obtenir une variété dont les caractéristiques correspondent au segment de marché ciblé par l'obtenteur.
Ceci explique le délai entre la mise à disposition des géniteurs et l'apparition des premières variétés porteuses des résistances qui en sont issues (Figure 1 p. 26). Depuis 2008 les six premières variétés résultant de ces recherches sont inscrites au catalogue.
Déterminants génétiques de la résistance
Des gènes dans les espèces sources
Les travaux conduits par l'UMR Igepp en lien étroit avec différents partenaires, dont l'équipe « solanacées à graines » de l'UGAFL, à l'Inra d'Avignon et la FN3PT (Fédération nationale des producteurs de plants de pomme de terre), ont permis de localiser plus ou moins finement les gènes de résistance aux parasites dans le génome de certaines des espèces sources.
Nématodes : nombre limité de QRL
Pour la résistance aux nématodes (Globodera et Meloidogyne), quelle que soit l'origine de la résistance, le nombre de zones chromosomiques impliquées dans le caractère de résistance (ou QRL pour quantitative resistance locus) est relativement limité : deux ou trois QRL ont pu être identifiés suivant les espèces.
Un QRL majeur pour la résistance à G. pallida est retrouvé de façon systématique sur le chromosome V, et explique une grande part de la variation du caractère alors qu'un ou deux QRL mineurs sont situés sur d'autres chromosomes.
Mildiou : plus de vingt gènes de type R
Dans le cas de la résistance au mildiou, le déterminisme génétique de la résistance est plus complexe. Des QRL à effet majeur correspondant à des gènes de type R conduisant à un blocage très efficace du développement du pathogène sont retrouvés.
Dans la littérature, plus de vingt gènes R ont été identifiés chez des espèces apparentées à la pomme de terre. Certains ont été introduits dans des variétés (gènes issus de S. demissum ou S. bulbocastanum). D'autres sont en cours d'exploitation, soit en utilisant la voie des croisements sexués, ce qui est le cas en France ou en Pologne, soit dans le cadre de travaux de cisgénèse conduits notamment aux Pays-Bas, qui consistent à introduire par génie génétique la séquence correspondant au gène de résistance.
Mais aussi la résistance partielle
Les chercheurs de l'Igepp s'intéressent particulièrement à une autre forme de résistance qui est sous contrôle polygénique : la résistance partielle. Pour des variétés porteuses de ce type de résistance, le développement de la maladie se montre beaucoup plus lent que sur les variétés sensibles.
Des travaux de cartographie ont permis de mettre en évidence de nombreux QRL répartis sur l'ensemble des chromosomes, chacun expliquant une faible part (inférieure à 20 %) de la variation du caractère. Certains génotypes peuvent combiner résistance totale (gènes R) et résistance partielle.
Que savons-nous des mécanismes de la résistance et de la durabilité de celle-ci ?
Des mécanismes hypothétiques
Si nous avons désormais des idées un peu plus précises sur les déterminants génétiques de la résistance à différents pathogènes, les mécanismes de défense chez la pomme de terre restent encore hypothétiques, en particulier pour ce qui est des résistances polygéniques plus complexes que les résistances mono ou oligogéniques.
Contre des nématodes ou des virus, certaines résistances sont durables
Les travaux de cartographie fine conduits sur S. sparsipilum, source de résistance à G. pallida, ont montré que le QRL majeur situé sur le chromosome V (nommé GpaVspl) pourrait être un gène de type R comme ceux identifiés chez différentes espèces sauvages résistantes au mildiou.
Dans le cas de ces gènes, des mécanismes de reconnaissance entre la plante et le nématode puis des cascades de signaux conduisent à des réponses de défense rapides et efficaces. Certains de ces gènes sont aujourd'hui largement déployés dans la gamme variétale et ne sont pas contournés après plus de quarante ans d'utilisation (gène H1 conférant la résistance à G. rostochiensis, gène Ry issu de S. stoloniferum conférant la résistance extrême au virus PVY).
Le mildiou contourne plus ou moins rapidement les gènes R
Il n'en est pas de même pour les gènes R issus de S. demissum ou de S. bulbocastanum conférant la résistance au mildiou.
En effet, ceux-ci ont été plus ou moins rapidement contournés par Phytophthora infestans dont les capacités d'adaptation, de multiplication et de diffusion sont extrêmement importantes.
Nématodes : contournement d'un gène en fonction du contexte génétique
Nos travaux permettent d'apporter un éclairage nouveau sur ces problèmes de contournement. En effet, les recherches conduites en étroite collaboration entre les généticiens et les nématologistes de l'Igepp sur la résistance à G. pallida ont démontré que le QRL GpaV ne fonctionne pas indépendamment du fonds génétique dans lequel il se trouve. Ainsi la résistance issue de la source S. vernei peut être contournée par le nématode après huit cycles de multiplication de celui-ci sur certains génotypes, alors qu'elle reste efficace dans les mêmes conditions pour d'autres génotypes. La notion de contournement devient donc plus relative : il n'y a pas contournement d'un gène de résistance mais contournement d'un gène dans un contexte génétique donné.
Nématodes : effets additifs
Plus encore, il a été clairement démontré, dans le cas de la résistance à G. pallida, qu'il existe des effets additifs entre GpaVspl et le QRL à effet mineur porté par le chromosome XI. Ainsi pour la résistance issue de S. sparsipilum, des nécroses apparaissent au niveau des tissus racinaires, bloquant la pénétration des larves dans les racines lorsque les deux QRL sont présents simultanément dans la plante, alors que si GpaVspl est seul présent, on observe uniquement une masculinisation des nématodes.
Mildiou : résistance partielle et architecture
Dans le cas de la résistance partielle au mildiou, les mécanismes sont moins bien décrits pour l'instant. Un QRL de résistance sur tige a été identifié sur le chromosome X de S. spegazzinii. Sa présence conduit à un ralentissement significatif de la progression du parasite dans la tige des plantes. Il ne semble pas limiter la progression des nécroses sur feuilles.
Dans le cadre du projet ANR Archidémio, il a pu être démontré qu'une partie de la résistance partielle peut être expliquée par des effets d'architecture de la plante et du couvert. Dans certaines conditions, en effet, un décalage significatif dans l'apparition des premiers symptômes de la maladie est observé, avec des couverts plus aérés. Des gènes impliqués dans la résistance et l'architecture de la plante pourraient se trouver aux mêmes endroits du génome, ou bien interagir entre eux. Des études sont encore nécessaires pour mieux comprendre ces phénomènes.
Même si la résistance partielle résultant du fonctionnement de plusieurs gènes offre davantage de garantie de durabilité, il n'en demeure pas moins que le parasite reste capable de se développer sur la plante. Cette coexistence rend possible, à plus ou moins long terme, l'émergence et la diffusion de souches de plus en plus adaptées.
Comment accompagner le déploiement de résistances pour les rendre plus durables ?
Construire des résistances complexes
Les problèmes évoqués plus haut concernant la durabilité des résistances génétiques rendent indispensable l'accompagnement du déploiement des gènes de résistance au moment de passer de l'échelle du laboratoire à celle de la sélection, puis de la production. La construction de résistances génétiques complexes est devenue un objectif majeur de nos recherches et un enjeu essentiel pour la création variétale. Les gènes de type R ne peuvent plus être utilisés seuls mais doivent être associés à des QRL mineurs ou mis dans des fonds génétiques favorisant leur durabilité.
Le déploiement de résistances partielles polygéniques doit être favorisé, en particulier pour des pathogènes très polymorphes et ayant de grandes capacités d'adaptation, comme P. infestans.
Ainsi, les géniteurs améliorés proposés par l'Inra aux obtenteurs français vont désormais résulter de croisements multiples permettant de cumuler les allèles favorables de différentes origines pour résister durablement à un ou plusieurs parasites simultanément.
Intérêt des marqueurs moléculaires
Le développement en parallèle et la mise à disposition de marqueurs moléculaires permettent, lorsque les allèles favorables sont bien identifiés, de s'assurer de leur présence aux étapes successives de sélection du matériel végétal (Encadré 4).
Conjuguer lutte génétique et autres moyens disponibles
Il convient aussi de conjuguer la lutte génétique avec toutes les autres méthodes de lutte disponibles (culturale, prophylactique, chimique) afin de limiter au maximum les risques de contournement et/ou d'adaptation. Des travaux expérimentaux et des efforts de modélisation sont réalisés pour combiner les différents leviers de lutte de façon optimale en tenant compte de la biologie des bioagresseurs.
Ils ont montré que l'usage en alternance dans le temps et l'espace (à l'échelle de la parcelle avec les mélanges variétaux, et aussi à celle de la région agricole) de différentes sources de résistance devrait permettre de limiter les risques d'apparition et de diffusion de variants plus adaptés. Des travaux doivent être poursuivis en ce sens. La lutte chimique apparaît toujours dans ces conditions comme un levier indispensable pour assurer une production suffisante, notamment pour lutter contre le mildiou.
Utiles OAD
Des outils d'aide à la décision (OAD) comme Mileos, en France, ont été développés pour permettre aux agriculteurs de mieux positionner leurs traitements et de choisir les substances actives adaptées aux conditions climatiques et de culture.
Des efforts ont été faits pour mieux prendre en compte la quantité d'inoculum primaire disponible au moment du démarrage de l'épidémie, mais il reste à intégrer ces éléments dans l'OAD et faire également évoluer celui-ci pour mieux prendre en compte la résistance variétale (niveau et type) dans la décision de traitement chimique.
Quelles sont les prochaines étapes ?
Six variétés pionnières et plus de cinq cents géniteurs pour en créer d'autres
Coquine, Cephora, Passion, Iledher, Malou et Stronga sont les variétés pionnières qui apparaissent aujourd'hui à l'issue d'un travail de recherche collaboratif entre l'Inra et les obtenteurs français initié depuis plus de quarante ans.
Entre 1995 et 2014, plus de cinq cents géniteurs de résistance ont été obtenus à l'Inra et proposés aux obtenteurs. Ceux-ci sont désormais en mesure d'exploiter cette grande diversité génétique pour créer des variétés résistantes aux stress biotiques et adaptées aux contraintes des marchés.
L'évolution récente des règles d'inscription au catalogue français devrait dans un proche avenir conduire à une augmentation progressive du niveau de résistance des variétés qui y sont inscrites. La contribution du matériel original provenant de l'Inra ainsi que la SAM (sélection assistée par marqueurs) devraient aider les sélectionneurs à répondre à ce renforcement des exigences.
Des recherches à poursuivre
Les recherches doivent toutefois être poursuivies pour mieux exploiter les nombreuses sources de résistance identifiées au sein du CRB BrACySol, notamment en essayant de mieux comprendre et exploiter les possibilités de recombinaison entre le génome de l'espèce cultivée et celui des espèces sauvages apparentées.
L'effort de modélisation devra être intensifié pour aider à l'amélioration des OAD par la prise en compte du déploiement de ce matériel végétal nouveau dans les stratégies de luttes combinées.
Sur le plan génétique, le développement de stratégies plus larges de génétique d'association en utilisant les collections du CRB BrACySol ou celles des obtenteurs devrait permettre d'amplifier les travaux d'identification et de localisation de déterminants génétiques impliqués dans des résistances durables, ce qui rendra la SAM de plus en plus opérationnelle.
Le cumul de résistances complexes contre plusieurs bioagresseurs, ainsi que la prise en compte des effets combinés de stress biotiques et abiotiques devraient, à terme, permettre la sélection de pommes de terre adaptées à des systèmes de culture plus économes en intrants et à l'agriculture biologique.
La recette pour assurer le succès des nouvelles variétés est particulièrement complexe. Elle fait intervenir des acteurs différents et nécessite des prises de risques bien au-delà de l'étape de l'inscription.
Les critères de résistance aux bioagresseurs doivent être considérés par les utilisateurs et prescripteurs au même niveau que les autres caractères, or les atouts qu'ils confèrent ne sont pas directement visibles. La volonté des différents acteurs de la filière et l'incitation des pouvoirs publics sont donc indispensables à la valorisation de ces variétés dans des itinéraires techniques adaptés.
1 – Le mildiou : ce qu'il faut savoir

Champ d'essai de l'Inra, à Ploudaniel, non traité contre le mildiou. Les différences de sensibilité entre géniteurs sont bien visibles. Photo : Inra
Le mildiou de la pomme de terre, provoqué par l'oomycète Phytophthora infestans, originaire du Mexique, reste, depuis l'introduction du parasite en Europe en 1845, le principal facteur limitant de la culture au niveau mondial.
Dès que les conditions de température et d'hygrométrie lui sont favorables, la maladie se déclenche. En l'absence de moyen de lutte approprié, elle connaît un développement extrêmement rapide qui peut conduire à la destruction totale du feuillage en moins d'une semaine.
La photographie présente un champ d'essais de l'Inra à Ploudaniel (zéro fongicide). L'effet des sensibilités variétales est nettement visible : les symptômes sur certaines variétés (ex. : Bintje) sont très marqués et d'autres génotypes restent indemnes (feuillage vert). Toutes les parties de la plante sont attaquées, si bien que la récolte peut se trouver réduite à néant si l'attaque a lieu avant la formation et le remplissage des tubercules.
Si l'attaque est plus tardive, la destruction du feuillage mais également des dégâts importants sur tubercules apparaissent. Il faut noter que la résistance au mildiou du tubercule est complètement indépendante de celle du feuillage.
Les programmes de sélection pour la résistance au mildiou, basés sur l'introduction de gènes de résistance monogénique issus en particulier d'espèces sauvages comme Solanum demissum ou S. bulbocastanum, ont montré des limites du fait de la grande variabilité du pathogène et de l'existence d'isolats capables de contourner ces résistances. L'exploitation de résistances partielles polygéniques offre de meilleurs espoirs de durabilité.
D'autres moyens de lutte peuvent être utilisés : lutte culturale et sanitaire (élimination des repousses et des tas de déchets, plantation de tubercules sains), lutte chimique avec des produits de contact (sels de cuivre ou mancozèbe), des matières actives pénétrantes ou translaminaires utilisées seules ou en association pour obtenir des effets curatifs. Un OAD (outil d'aide à la décision), Miléos, a été développé en France afin d'aider les agriculteurs dans le positionnement des traitements et le choix des molécules les plus adaptées pour limiter les risques de développement épidémiques... et d'apparition d'isolats résistants.
2 – Quels traitements employer face aux nématodes « à kyste » ?

Les petites boules blanches sur ces racines de pomme de terre sont des femelles du nématode à kyste Globodera pallida. Photo : Inra
Les nématodes à kyste Globodera rostochiensis et G. pallida sont originaires du Pérou et ont été introduits en Europe au XIXe siècle sur des tubercules de pomme de terre.
Ce sont des parasites obligatoires qui attaquent spécifiquement les solanacées.
Un kyste peut contenir plus de cinq cents œufs et survivre dans le sol plus de dix ans.
En présence d'exsudats racinaires, les larves sont libérées, colonisent les tissus racinaires et évoluent préférentiellement en femelle en présence d'une variété sensible (la photographie représente des femelles de couleur blanche sur les racines d'une telle variété).
Les œufs se développent à l'intérieur des femelles qui s'enkystent et se détachent des racines à maturité.
En fonction de la densité de kystes présents dans une parcelle, les dégâts peuvent être d'ordre quantitatif (jusqu'à 70 % de perte de rendement) ou qualitatif (les piqûres déprécient la qualité des tubercules).
Au niveau mondial, les nématodes sont classés parmi les parasites de quarantaine. De ce fait, la commercialisation de plants infestés est interdite et les méthodes de lutte sont obligatoires (directive européenne 2007/33/CE et JORF du 1er juillet 2010).
Le traitement des sols par des nématicides représente l'une des méthodes de lutte. Il s'avère efficace mais peu respectueux de l'environnement. Actuellement, il n'existe plus que trois substances actives homologuées : l'oxamyl, le fosthiazate et le dazomet.
Les méthodes alternatives, quand elles sont applicables, ne sont pas toujours satisfaisantes. Ainsi, l'efficacité de la solarisation dépend des conditions climatiques. L'utilisation des plantes-pièges en interculture permet de réduire les populations de nématodes dans le sol.
La lutte génétique trouve sa place dans ce contexte. Il s'agit d'exploiter des génotypes empêchant la multiplication des nématodes. La recherche s'orientera donc vers des génotypes qui conduisent le développement du nématode vers un stade mâle (« masculinisation ») ou qui bloquent la pénétration ou le développement des nématodes dans les racines.
Concernant la résistance à G. rostochiensis, un gène majeur a été identifié chez Solanum tuberosum ssp. andigena (espèce très proche de la pomme de terre cultivée en Europe). Ce gène H1 a été largement introduit dans les variétés modernes.
En France, le déploiement de variétés résistantes à G. rostochiensis avait eu pour conséquence le remplacement de cette espèce par sa « cousine » G. pallida, qui n'était jusqu'alors pas prise en compte dans les schémas de sélection.
Fig. 1 : Différentes étapes ayant précédé l'inscription de six variétés résistantes au catalogue français par les quatre sélectionneurs français réunis au sein de l'ACVNPT
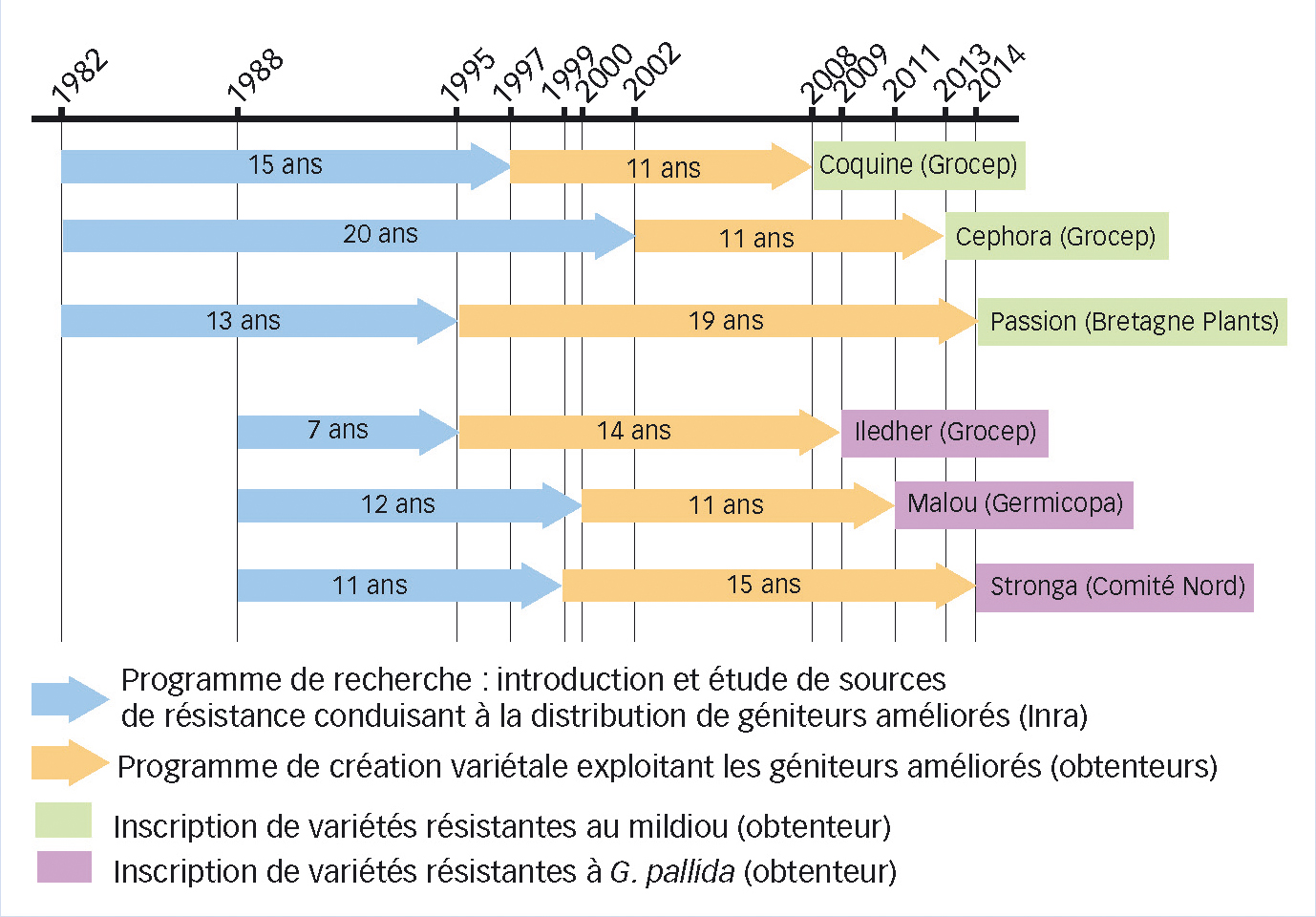
Le temps nécessaire à l'obtention d'une variété sera plus ou moins long (ici, de 21 à 32 ans) selon, notamment, la source de résistance introduite (espèce sauvage ou cultivée).
3 – Comment identifier une résistance dans les collections ?
Le Centre de ressources biologiques BrACySol, maintenu par l'UMR Igepp sur le site Inra de Ploudaniel, entretient une collection importante de solanacées tubéreuses par voie végétative (au champ, en serre ou in vitro).
Cette collection, qui comporte plus de 12 000 accessions, comprend, entre autres, plus de 1 000 clones appartenant à une trentaine d'espèces apparentées à la pomme de terre, originaires d'Amérique du Sud. Ces clones ont été évalués pour leur résistance à différents bioagresseurs, dont le mildiou de la pomme de terre provoqué par l'oomycète Phytophthora infestans, le nématode à kyste Globodera pallida, et quelques nématodes à galles (Meloidogyne sp.).
Concernant la résistance au mildiou, l'évaluation est réalisée par l'UMR Igepp sur le site de Ploudaniel où les conditions climatiques sont favorables à l'expression de la maladie.
Différents tests sont mis en œuvre. Dans le cadre d'expérimentations en pots sur plateforme extérieure, la vitesse de destruction du feuillage des différents génotypes est mesurée en conditions naturelles de contamination.
Ce test permet d'évaluer le niveau et le type de résistance (monogénique totale ou polygénique partielle) au mildiou du feuillage.
D'autres tests réalisés en serre permettent de mesurer la progression du parasite dans les tiges après inoculation de plantes entières avec des souches bien caractérisées de mildiou.
Ces tests conduisent à évaluer des composantes de la résistance partielle sur tige (c'est-à-dire la réceptivité de la plante, la capacité à mettre en place une réaction de défense, l'aptitude à maintenir un mécanisme de résistance stable ainsi que le niveau global de résistance).
Ainsi, quatorze sources de résistance ont été identifiées, dont certaines sont originales, comme S. polytrichon ou S. trifidum.
Pour les nématodes étudiés qui sont pour la plupart des parasites de quarantaine, l'évaluation pour la résistance est mise en œuvre par l'UMR Igepp sur le site du Rheu.
Les tests, réalisés en pots en serre confinée (niveau S2) ou sur plantules in vitro, ont permis d'évaluer la capacité de développement des nématodes dans les génotypes après inoculation avec des larves de populations bien caractérisées. Ainsi, trois sources de résistance à Globodera pallida (Solanum vernei, S. sparsipilum et S. spegazzinii), et trois sources de résistance aux Meloïdogyne sp. (S. sparsipilum, S. polytrichon et S. schenckii) ont été identifiées.
4 – La SAM, sélection assistée par marqueurs
La sélection de nouvelles variétés de pomme de terre prend du temps. Il faut compter environ dix ans entre le croisement et la mise à l'inscription des clones prometteurs au catalogue. Dix années d'une sélection familiale, individuelle et multilocale sur une trentaine de critères : agronomiques (rendement, précocité, teneur en matière sèche...), technologiques (noircissement après cuisson, fritabilité, sucrage à basse température...) et de résistance aux bioagresseurs (mildiou, nématodes, virus, etc.).
La sélection assistée par marqueurs (SAM) consiste à utiliser précocement, au laboratoire, des marqueurs ADN prédictifs de la valeur des clones pour certains caractères. Employée en complément de la sélection conventionnelle, la SAM permet d'aller plus vite et de gagner en efficacité en se concentrant sur les clones les plus intéressants pour l'expérimentation au champ. Elle permet également de réaliser des tests qu'il serait impossible de faire en sélection classique, par exemple l'identification d'hybrides possédant des facteurs génétiques améliorants. L'efficacité de la SAM repose sur la fiabilité des marqueurs. Leur mise au point est donc primordiale.
La résistance aux bioagresseurs est, pour le moment, le champ d'utilisation majeur de la SAM pour l'amélioration de la pomme de terre. Partout dans le monde, des marqueurs pour différents gènes de résistance aux nématodes à kyste et aux virus sont utilisés. Les sources de résistance sont assez peu diversifiées et, fort heureusement, non contournées. Le travail d'identification d'une source de résistance originale au nématode G. pallida réalisé à l'Inra a permis la mise à disposition des sélectionneurs français de géniteurs portant la résistance et des marqueurs associés.
Avec l'évolution rapide des méthodes et technologies dans le domaine de la génomique, il est maintenant possible d'avoir une vision globale du génome d'un hybride.
À moyen terme, l'accès par la SAM aux autres critères de sélection sera possible, notamment à ceux dont la sélection est difficile pour d'autres raisons que celles évoquées précédemment comme une forte interaction entre génotype et environnement, ou encore une hérédité compliquée fortement polygénique. La majorité des caractères suivis par les sélectionneurs, en somme...

