
 Imprimer
Imprimer")
Tableau 1 : Fréquence des allèles virulents vis-à-vis du gène Rlm7 dans les populations françaises de phoma (Leptosphaeria maculans)

Tableau 2 : Type et fréquence des événements mutationnels dans le gène AvrLm7, responsables de l'acquisition de la virulence vis-à-vis des variétés Rlm7

Les souches virulentes seraient générées par reproduction sexuée sur les résidus. Photo : T. Rouxel - Inra
La nécrose du collet du colza, ou phoma du colza, est l'une des principales maladies cryptogamiques de cette culture (Fitt et al., 2006). Face à cette maladie, la résistance variétale est un outil... à utiliser avec vigilance. En voici la preuve, à propos du gène Rlm7.
Des histoires de génétique
Régression de l'impact du phoma grâce à la génétique
Ces dix dernières années, la maladie est restée discrète. Cette situation est liée à des conditions météorologiques peu favorables aux épidémies, mais surtout à l'amélioration génétique des variétés de colza pour leur résistance à la maladie.
En effet, le comportement variétal du colza vis-à-vis du phoma est un des critères principaux pris en compte par les sélectionneurs. Par ailleurs, le CTPS a mis en place de longue date un règlement qui promeut l'inscription de variétés résistantes et élimine les variétés trop sensibles.
Résistance variétale : les deux types
Deux types de résistance au phoma sont généralement décrits (Delourme et al., 2006). - d'une part les résistances quantitatives, efficaces vis-à-vis de toutes les souches de l'agent pathogène, mais n'apportant qu'un niveau partiel de résistance (les plantes peuvent être infectées, mais l'intensité de la nécrose au collet reste réduite et impacte peu le rendement). Le niveau de résistance quantitative des variétés est en constante augmentation.
- d'autre part, les résistances contrôlées par des gènes majeurs de résistance, qui bloquent l'infection primaire donc apportent une protection totale, mais uniquement vis-à-vis de la fraction de la population pathogène portant les allèles avirulents des gènes d'avirulence correspondants (Encadré 1) (Balesdent et al., 2002).
Le gène Rlm1, grandeur et décadence
Le gène de résistance spécifique Rlm1 avait été introduit dans les variétés de colza au début des années 1990, dans un contexte où plus de 90 % des souches de phoma portaient l'allèle avirulent du gène correspondant, AvrLm1 (Rouxel et al., 2003).
Ce gène s'est révélé très efficace. Il a donc été très utilisé, avec 20 % des surfaces de colza françaises portant Rlm1 en 1995 et 40-45 % en 1999 (Rouxel et al., 2013).
Ce gène a ainsi exercé une pression très forte sur le champignon. Celle-ci a conduit à une sélection rapide de souches virulentes dans les populations de phoma. Cela s'est traduit par un contournement de la résistance en quelques années.
Ce phénomène est une illustration typique d'un cycle de « boom-and-bust » et du potentiel adaptatif des parasites aériens en grande culture (Rouxel et al., 2003).
En 2004, arrivée de Rlm7
Depuis cet épisode de contournement, un nouveau gène de résistance spécifique, Rlm7, a été introduit dans des hybrides de colza commercialisés à partir de 2004, dans un contexte où 99,9 % des souches de phoma étaient avirulentes vis-à-vis de Rlm7 (Balesdent et al., 2006). Ces hybrides ont donc connu un très large succès.
Les enquêtes du Cetiom évaluaient leurs parts de marché à 27 % en 2009. Aujourd'hui ces hybrides occupent environ 50 % des surfaces au niveau national et peuvent atteindre localement 70 %.
Dans cet article, nous ferons un bilan des études de populations de phoma réalisées ces quinze dernières années, pour illustrer l'état actuel du contournement de Rlm7 à l'échelle du territoire, en comparaison avec le contournement passé de Rlm1.
Analyse des populations de phoma
À l'échelle nationale, trois campagnes à quelques années d'intervalle
Trois grandes campagnes d'analyses de populations de phoma ont été menées à l'échelle du territoire : en 2000-2001, en 2009-2010 et en 2013 (Tableau 1).
Pour chaque campagne, une variété sans gène de résistance (Drakkar) ou avec un gène de résistance déjà contourné (Falcon) est semée.
À l'automne, des feuilles portant des macules typiques de phoma sont prélevées, chacune sur une plante différente. L'effectif visé est de 80 à 100 souches par site.
Des isolements monopycnidiens sont alors réalisés sous loupe binoculaire à partir de ces macules, et les souches sont ensuite caractérisées soit par inoculation de suspensions de spores sur une gamme de variétés différentielles (Balesdent et al., 2006), soit à l'aide d'outils moléculaires (Carpezat et al., 2014) (Encadré 2).
Résultats : une évolution indéniable
En 2000-2001, une seule souche analysée sur plus de 1 700 était virulente vis-à-vis de Rlm7.
Dix ans plus tard, la fréquence des souches virulentes atteint 4 %. Leur présence est détectée dans la moitié des sites échantillonnés, avec de très grandes variations suivant les sites (0 % à 22,5 %, Figure 1).
À l'automne 2013, la moyenne nationale se situe autour de 19 % (Tableau 1), et des souches virulentes sont détectées dans tous les sites sauf Rennes, à des fréquences très variables (4 à 45 %) suivant les sites.
Au plan régional, les quatre sites du centre et de la Beauce montrent des fréquences de virulence proches (37,6 à 45 %), suggérant un réel contournement de Rlm7 dans cette zone (Figure 1). En Normandie et dans l'est, les fréquences sont inférieures à 10 %. Pour le reste des sites les fréquences sont comprises entre 12 % et 17 %.
Suivi annuel en région Centre depuis 2004
Dans une zone de la région Centre fréquemment atteinte par le phoma, à l'échelle de quelques communes, le Cetiom a réalisé un suivi annuel depuis 2004.
Si aucun individu virulent n'a été détecté jusqu'à l'automne 2009 sur les variétés sans le gène Rlm7, quelques individus virulents ont été détectés lors des automnes 2010 et 2011 (fréquence inférieure à 3 %).
Ensuite, on note une explosion des individus virulents : 49 % et 45 % à l'automne 2012 et 2013, respectivement.
Pratiques culturales et contournement
Étude de cas extrêmes sur deux sites en Île-de-France
Pour tenter de comprendre les facteurs favorisant la sélection et le succès des souches virulentes, un essai a été conduit sur plusieurs années à l'Inra. Le travail a été réalisé sur les sites expérimentaux de Versailles et de Grignon, distants d'une vingtaine de kilomètres.
Cette expérimentation visait à mesurer l'impact de pratiques culturales favorables à la maladie sur la vitesse de sélection des souches virulentes. Sur ces deux sites, les variétés Rlm7 Roxet (2004-2005) puis Exagone (2005-2008) ont été cultivées.
Sur le site de Versailles les cultures ont été implantées avec une rotation colza-céréales, un intervalle supérieur à trois ans entre deux cultures de colza et un labour après colza pour enfouir les résidus.
Sur le site de Grignon, les variétés Rlm7 ont été semées sur la même parcelle, et seul un travail simplifié du sol est réalisé entre deux cultures.
Ces deux sites représentent donc des situations extrêmes, soit très prudentes en terme de contrôle de la maladie (Versailles) soit très risquées (Grignon).
Dans les deux sites, une bande de 10 mètres de large, contiguë aux parcelles portant Rlm7, est semée avec une variété « piège », ne portant pas Rlm7, sur laquelle sont réalisés chaque année des échantillonnages de populations de phoma à partir de macules foliaires développées à l'automne.
Au cours de ce travail, 1 309 souches de phoma ont été ainsi isolées et phénotypées pour leur virulence vis-à-vis de Rlm7. Les populations locales avaient été caractérisées sur les deux sites avant l'expérimentation. Enfin, des populations locales ont été échantillonnées à proximité de la parcelle expérimentale de Grignon, pendant et quelques années après la fin de l'expérience (2006, 2009, 2011, 2012 et 2013).
Résultat : oui, les pratiques culturales ont un effet sur le contournement !
Cet essai a révélé une différence très significative entre les évolutions des fréquences de souches virulentes sur les deux sites :
- à Versailles, les souches virulentes restent indétectables pendant toute la durée de l'expérimentation.
- à Grignon, la fréquence de souches virulentes atteint 9,3 % dès la seconde année de pratiques culturales « risquées », soit en 2006 ; elle triple en 2007 (Figure 2).
Cette évolution des fréquences de virulence reste très localisée à la parcelle expérimentale elle-même. En effet, aucune souche virulente n'est détectée en 2006 dans une parcelle de colza située à 500 m de la parcelle expérimentale (Figure 2).
Effet du passage à des pratiques « prudentes »
Fin 2008, la parcelle expérimentale de Grignon a été labourée après récolte du colza. Les fréquences de virulence ont été suivies sur des parcelles de colza proches de 2009 à 2013. En 2009, le pourcentage de souches virulentes est très faible (1,25 %). Ensuite, les fréquences de virulences vont progresser au rythme de l'évolution à l'échelle nationale (losanges jaunes Figure 2).
Quel est le moteur de la génération d'isolats virulents ?
Analyse moléculaire des souches virulentes
Pour identifier les événements moléculaires responsables de l'acquisition de la virulence vis-à-vis de Rlm7, 769 souches virulentes issues de la parcelle expérimentale de Grignon ont été caractérisées. L'analyse moléculaire a été réalisée par amplification PCR du gène AvrLm7 et séquençage de l'allèle si le gène était amplifié (Daverdin et al., 2012).
Ce travail a montré l'existence d'un grand nombre d'événements moléculaires différents responsables de l'acquisition de la virulence (Tableau 2). De plus, pour chaque type d'événement, un très grand nombre d'allèles différents est observé. Cette diversité a été observée sur une population isolée de l'unique parcelle expérimentale de Grignon, ceci dès la première année d'étude.
Rôle de la reproduction sexuée du champignon
Parmi ceux-ci, la délétion totale ou partielle du gène, l'inactivation par mutations multiples dues au mécanisme de RIP (Repeat-induced point mutation) (Fudal et al., 2009) et l'insertion d'éléments transposables dans la séquence du gène sont générés spontanément au cours de la méiose donc de la reproduction sexuée. Ces événements totalisent à eux seuls 87,6 % des cas de virulence observés (Tableau 2).
Il est donc suggéré que des souches virulentes peuvent être générées localement chaque année suite à la reproduction sexuée du champignon, laquelle a lieu sur les résidus de culture.
Les ascospores, produits de la méiose, constituent en effet l'inoculum primaire. Les souches virulentes ainsi générées risqueront d'autant plus d'être sélectionnées qu'une variété portant Rlm7 sera cultivée à proximité immédiate de la culture précédente.
Durabilité de Rlm7 : les quatre hypothèses
L'analyse de l'évolution des fréquences de virulence dans les populations de phoma en France suggère une durabilité potentielle plus élevée de la résistance Rlm7 que de Rlm1 : en trois ans d'usage de Rlm1, les souches virulentes vis-à-vis de ce gène ont constitué plus de 80 % de la population française, alors qu'après dix ans de Rlm7 la fréquence moyenne des souches virulentes reste inférieure à 25 % (Figure 3). Plusieurs hypothèses pourraient expliquer cela.
Hypothèse « suivi des conseils » ?
La première serait l'effet positif des conseils d'alternance variétale (alterner résistances quantitatives et résistances spécifiques) et de rotations évitant des retours trop fréquents du colza sur la même parcelle.
Néanmoins, en pratique, l'évolution tend plutôt vers le raccourcissement des rotations et l'augmentation des superficies cultivées avec du colza Rlm7 !
Rôle du climat : hypothèse recevable
Des conditions climatiques différentes, plus ou moins favorables à la maladie, ont pu davantage favoriser le contournement de Rlm1 que celui de Rlm7.
Effet « construction des variétés »
Une troisième hypothèse serait la construction des variétés Rlm7 commercialisées. En effet, il a été démontré que l'association d'une résistance spécifique avec un fort niveau de résistance quantitative accroît nettement la durabilité de la résistance spécifique (Brun et al., 2010). Des variétés Rlm7 pourraient être dans ce cas.
Tant que leur résistance Rlm7 n'est pas contournée, il est impossible d'évaluer leur résistance quantitative. Avec des sites approchant 50 % de souches virulentes en 2013, des informations objectives sur la composante quantitative des variétés Rlm7 devraient être disponibles à court terme.
Fréquence initiale des virulences
La quatrième hypothèse réside dans le niveau initial de fréquence des virulences lors du début de l'utilisation commerciale du gène de résistance.
Quand les premières variétés Rlm1 ont été introduites en France, les fréquences de souches virulentes avoisinaient 10 % et il n'a fallu que trois années pour constater une inversion des fréquences de virulence à l'échelle nationale (Figure 3). On peut supposer qu'il faut une période de temps assez longue pour que les fréquences de virulence atteignent un niveau décelable (dans le cas de Rlm7, pour passer de 0,06 % de virulentes en 2000 à environ 4 % en 2010), mais que le contournement s'accélère une fois atteint des fréquences avoisinant 5-10 % .
Le suivi du Cetiom en région Centre depuis 2004 appuie cette hypothèse.
Elle sera testée par suivi des populations sur les sites échantillonnés en 2013 durant deux années supplémentaires dans le cadre du projet « Dynamic-Rlm7 »(1).
Faire face à la nouvelle phase
Risque d'accélération
Les résultats menés dans le site-pilote du Cetiom, ainsi que l'observation à l'échelle du territoire de l'évolution entre 2009 et 2013 des fréquences des virulences, suggèrent que l'on pourrait être arrivé dans une phase de contournement accéléré.
En conditions expérimentales maximisant le succès des souches virulentes, l'évolution a été très rapide : de 0 à 36 % en trois ans.
Alternance variétale, rotation et destruction des résidus
Dans cet essai, les parcelles situées à quelques kilomètres et sur lesquelles des pratiques culturales optimales ont été appliquées (rotations > 3 ans, enfouissement des résidus par labour) ont été préservées de l'apparition de souches virulentes.
Il est donc crucial de préserver les régions dans lesquelles les fréquences de virulence sont encore faibles par un respect des délais de retour du colza, des règles de diversification et d'alternance des variétés et des pratiques culturales visant à la destruction des résidus de cultures.
En effet, ces derniers sont le lieu non seulement de la génération de l'inoculum de l'année suivante, mais aussi de la diversification allélique conduisant à la génération de souches virulentes.
Existe-t-il une cinquième hypothèse ?
Par ailleurs, un autre mécanisme a pu contribuer à ralentir l'apparition des souches virulentes vis-à-vis de Rlm7 : il s'agit de la relation entre l'acquisition de la virulence avrLm7 et la résurgence du phénotype avirulent sur les variétés possédant Rlm3 (Daverdin, 2011).
Ce mécanisme et ses conséquences en termes de gestion des résistances sont développés dans l'article suivant.
(1) Projet 2015-2016, financé par le progr. ANR « Investissements d'avenir » (LabExBASC ; ANR-11-LABX-0034).
1 - Les interactions gène pour gène dans le modèle phoma-colza
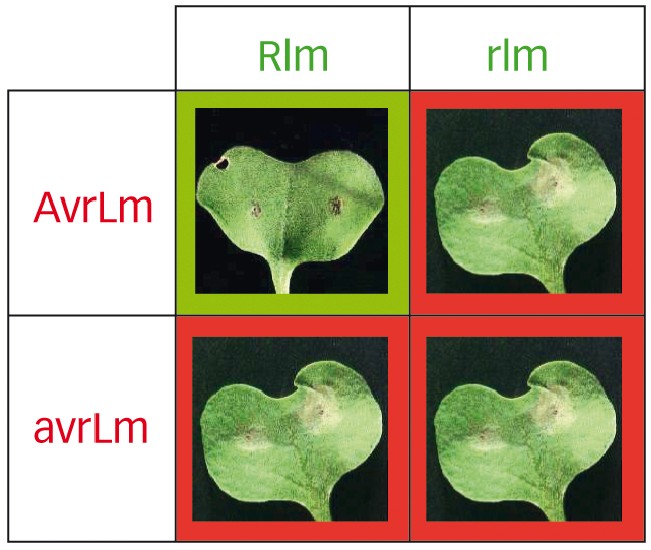
Lorsque la plante possède un gène de résistance spécifique Rlm, elle développe une réaction de résistance vis-à-vis des souches possédant l'allèle avirulent du gène d'avirulence correspondant, noté AvrLm (case verte).
Dans tous les autres cas, la plante ne détecte pas la présence de l'agent pathogène. Ce dernier peut alors induire les symptômes typiques de la maladie (macules foliaires, cases rouges).
2 - Méthode d'obtention et de caractérisation de populations de Leptosphaeria maculans

1/Découpage d'une macule de phoma typique sur feuille de colza, et mise en « chambre humide » pendant 48 heures.
2/Observation à la loupe des pycnides sporulantes (gouttelettes rosées), prélèvement à l'aide d'une aiguille stérile et dépôt des spores sur milieu de culture.
3/Purification et mise en culture des souches en boîte de Petri.
4/Phénotypage des souches par inoculation sur variétés différentielles puis observation à l'oeil nu (ici, la souche est virulente sur le génotype de colza représenté par le cotylédon de gauche cf. les macules, mais avirulente sur celui du cotylédon de droite).
5/Alternativement, caractérisation par analyse moléculaire, ici méthode HRM (Carpezat et al., 2014).
Fig. 1 : Localisation des sites analysés en 2009-2010 et 2013 et fréquence des souches virulentes (en rouge) vis-à-vis de Rlm7
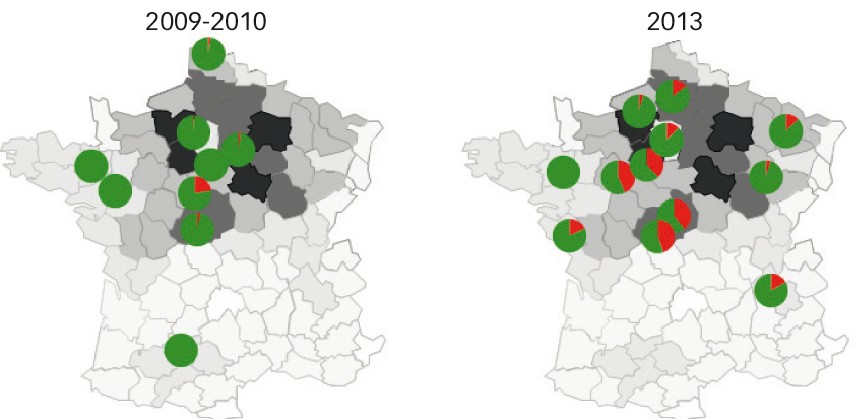
L'intensité du grisé traduit l'intensité de la culture de colza sur chaque département en 2013 (source : Geves).
Fig. 2 : Évolution des fréquences de virulence vis-à-vis de Rlm7 selon les pratiques culturales
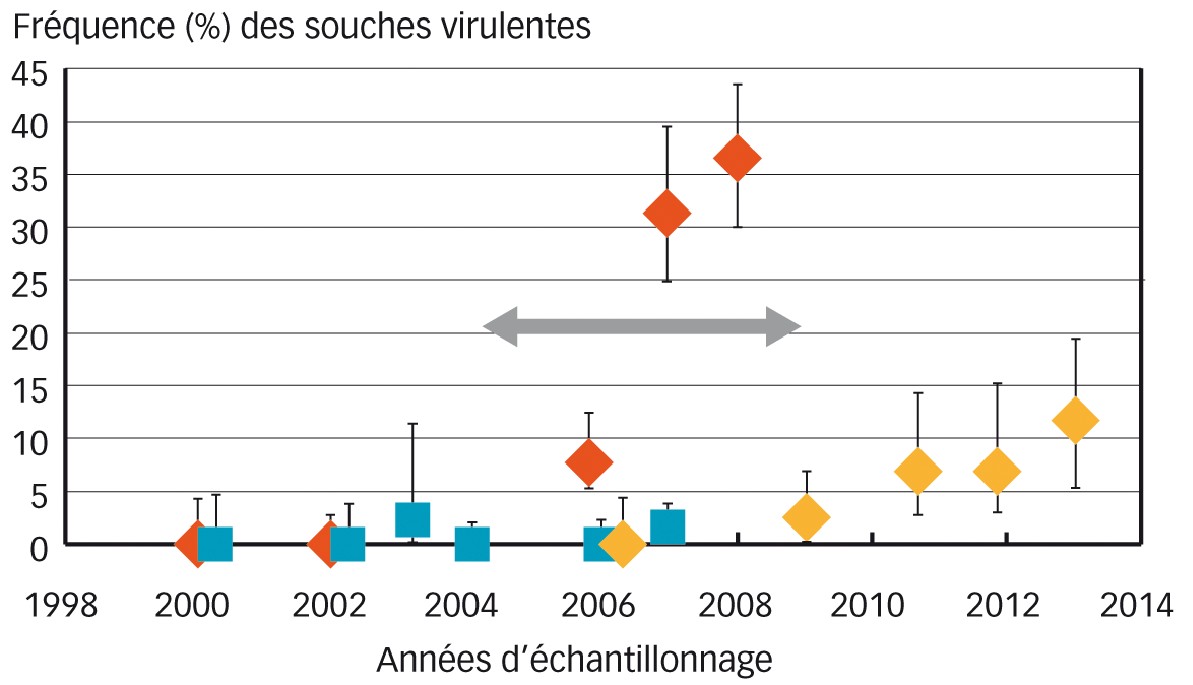
La fréquence de souches virulentes a été estimée sur des populations échantillonnées sur variétés pièges sans Rlm7.
Losanges : populations sur le site de Grignon ; les rouges correspondent à la population de la parcelle expérimentale soumise aux « pratiques culturales risquées » ; les jaunes correspondent aux populations prélevées sur le site (distance maximale à la parcelle expérimentale = 800 mètres) soumises à des pratiques standard (rotation, labour).
Carrés bleus : populations échantillonnées à Versailles (pratiques standard).
Barres verticales : intervalle de confiance exact calculé en fonction des effectifs analysés.
Flèche grise : période d'expérimentation « pratiques culturales risquées » sur Grignon.
Fig. 3 : Évolution comparée des fréquences des souches virulentes vis-à-vis de Rlm1 et Rlm7
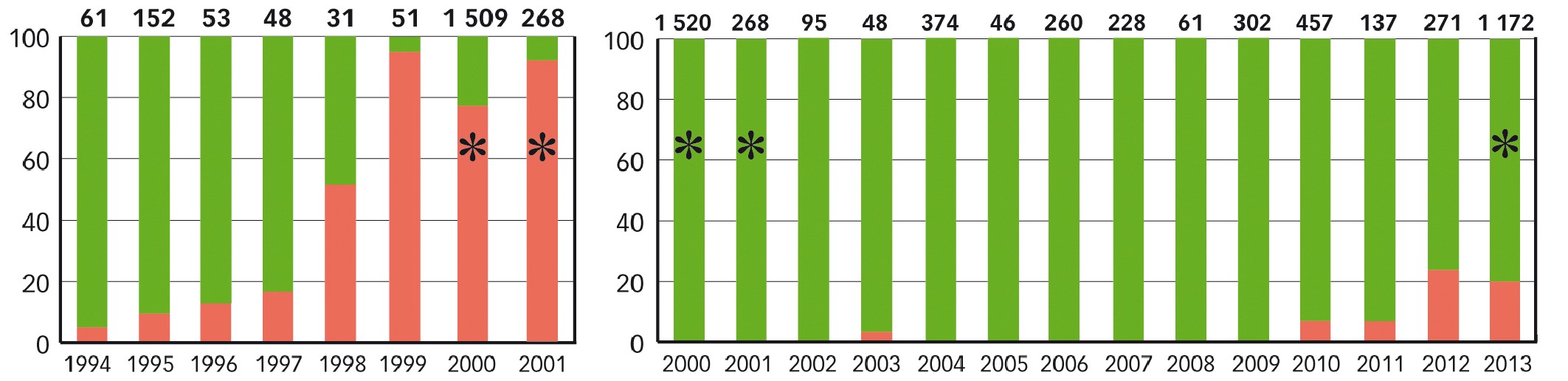
Les étoiles indiquent les échantillonnages nationaux. Les chiffres en gras représentent le nombre de souches analysées par année. Les barres rouges correspondent aux fréquences de souches virulentes ; diagramme de gauche, virulence sur Rlm1 ; à droite, virulence sur Rlm7.
SUMMARY
RESISTANCE TO CONTROL PHOMA ON OILSEED RAPE - In France, the major resistance gene Rlm7 has been deployed on a wide scale these last 10 years to control phoma (stem canker) on oilseed rape. It was successfull but, now, phoma leaf spots can be observed on some Rlm7 genotypes.
This fact suggests that the resistance could be overwhelmed.
This paper summarizes our current knowledge on virulent isolates toward Rlm7 in French phoma populations. The ways to increase Rlm7 durability are discussed.
KEYWORDS - Oilseed rape, France, phoma (stem canker, Phoma lingam, Leptosphaeria maculans), resistance, virulente isolates, Rlm7 genotypes, overwhelmed, durability.
REMERCIEMENTS
Ce travail a bénéficié d'un soutien financier du CTPS (Projet Evolep C3-2008-Phoma, Projet Icoscop 2012-2015), de l'ANR (Projet Gester) et du métaprogramme Inra SMaCH (Projet K-masstec). Les auteurs remercient C. Montagnier (Unité expérimentale Inra-Grignon) pour la gestion des parcelles expérimentales de Versailles et Grignon.

