
 Imprimer
Imprimer
1. Symptômes de flavescence dorée sur baroque. Ce cépage béarnais étant blanc, la flavescence y est dorée. Photos : J. Larrue - Inra Dijon

2. Symptômes sur grenache. Ce cépage étant noir, les feuilles atteintes rougissent. Photos : J. Larrue - Inra Dijon

3. Adulte de Scaphoideus titanus, vecteur de la maladie. Cette cicadelle américaine s'est remarquablement adaptée aux vignes européennes. Photo : Inra

4. À gauche, cep de chardonnay exprimant la flavescence dorée : les grappes se sont formées mais ont flétri ; par ailleurs, les feuilles montrent le jaunissement caractéristique et les rameaux restent verts. À droite, cep de chardonnay semblant sain... mais qui est peut-être en incubation ! Photo : J. Larrue - Inra Dijon

5. Phytoplasmes dans une coupe longitudinale de tube criblé de vigne malade. Observation au microscope électronique à transmission. Photo : J. Lherminier - Inra Dijon
La flavescence dorée appartient à un groupe de maladies appelées jaunisses de la vigne, connues dans tous les pays viticoles et se ressemblant par leur symptomatologie. La flavescence dorée (photos 1 et 2) fut la première jaunisse de la vigne jamais décrite. Elle est aussi particulièrement nuisible. Voyons ici pourquoi et comment.
Historique
Les jaunisses de la vigne sont étudiées depuis la première découverte
La flavescence dorée apparut en Gascogne en 1955. Elle fut d'abord attribuée à un désordre physiologique. Son caractère infectieux fut démontré lorsque son vecteur, la cicadelle Scaphoideus titanus Ball (photo 3), fut identifié (Schvester et al., 1963).
On a d'abord pensé que l'agent infectieux était un virus (Caudwell, 1964), jusqu'à ce que la découverte, en 1967, des « MLO » (mycoplasma-like organisms, appelés plus tard phytoplasmes) chez les plantes, oriente la recherche vers ces nouveaux agents pathogènes.
Ils furent alors observés dans le corps de spécimens de S. titanus vecteurs de la flavescence dorée et dans les vignes malades inoculées par ces derniers (Caudwell et al., 1971). Pendant les décennies suivantes, des maladies ressemblant à la flavescence dorée furent décrites dans d'autres pays (Gärtel, 1965 ; Pearson et al., 1985).
Il fut décidé en 1992 de réserver le nom de flavescence dorée à la seule maladie associée à l'activité vectrice de la cicadelle S. titanus (Bovey & Martelli, 1992).
À partir des années 1990, le développement de méthodes de biologie moléculaire de plus en plus fines conduisit à l'identification des phytoplasmes associés aux différentes jaunisses de la vigne.
Les données d'épidémiologie qui s'appuyaient sur les observations biologiques et agronomiques se sont dès lors enrichies des comparaisons de souches et d'isolats, effectuées par divers laboratoires européens (voir article p. 21).
Les récents épisodes de contaminations découvertes dans des vignobles prestigieux et les contraintes de lutte contre cette maladie de quarantaine ont donné un retentissement médiatique à cette maladie. Le grand public, et même des viticulteurs, l'ont découverte à cette occasion.
Pourtant, la flavescence dorée a été décrite il y a près de soixante ans (Caudwell, 1957). Les recherches, très fastidieuses au départ, ont été conduites dès cette date, mariant sans cesse les observations de terrain et les tentatives de reproduction au laboratoire pour parvenir aux connaissances actuelles et aux outils moléculaires encore en développement.
Épidémies en France
Au cours des années 1960, la flavescence dorée détruisit une partie des vignobles de la Chalosse et de l'Armagnac. Une deuxième épidémie apparut en Corse en 1970, tandis qu'un mitage des zones viticoles du Sud-Ouest et du Sud était observé.
Une troisième épidémie se déclara au début des années 1980 à l'ouest du département de l'Aude pour envahir progressivement toute la région Languedoc-Roussillon.
D'autres foyers importants se manifestèrent en Aquitaine, Charente (Roure, 2000), Rhône-Alpes et PACA.
En 2006, 2 241 communes viticoles (source SDQPV) étaient soumises aux mesures de lutte obligatoire. Cependant, la situation des vignobles derrière le front de progression est très contrastée. Bien que l'assainissement des parcelles atteintes puisse être réalisé progressivement dans les zones de lutte, des foyers ponctuels peuvent malgré tout se déclarer au sein de vignobles stabilisés.
Ailleurs en Europe
En Italie, Vidano (1964) observa la présence de S. titanus en Italie du Nord.
En 1975, une maladie de type flavescence dorée fut décrite en Oltrepò Pavese-Lombardie et associée à la présence de S. titanus (Belli et al., 1973 ; Osler et al., 1975). Une controverse sur la présence effective de la flavescence dorée en Italie découla de la présence simultanée du bois noir (non transmis par S. titanus) dans les vignobles italiens, alors qu'aucun diagnostic différentiel n'était disponible.
Au début des années 1990, la progression rapide de la maladie suggéra fortement la présence, dans plusieurs régions italiennes, de flavescence dorée. Elle fut confirmée par les méthodes sérologiques et moléculaires tout juste élaborées (Osler et al., 1992 ; Daire et al., 1992, 1993 ; Bertaccini et al., 1995).
En Espagne, la flavescence dorée fut identifiée pour la première fois en 1996 au nord-est de la Catalogne (Batlle et al., 1997) puis à nouveau en 1999.
Plus tard, elle fut identifiée en Serbie (2002), en Suisse dans le canton du Tessin (2004), au Portugal (2005), en Slovénie (2008), et plus récemment en Hongrie et Roumanie.
La flavescence dorée a été décrétée organisme de quarantaine dans la Communauté européenne (directive CE n° 77/1993 modifiée 92/103). La déclaration de la maladie et le détail des mesures de lutte ont fait l'objet de plusieurs arrêtés ministériels en France (17 avril 1987 et 1er avril 1994 remplacés par celui du 9 juillet 2003, puis celui du 19 décembre 2013, modifié en septembre 2015) et en Italie (31 mai 2000).
Distribution géographique du vecteur et de la maladie
Origine et distribution de Scaphoideus titanus
L'espèce Scaphoideus titanus Ball, 1932, est une Deltocephalinae de la famille des Cicadellidae. Au sein du genre nord-américain Scaphoideus Uhler, 1889, elle est la seule espèce décrite en Europe (= S. littoralis Ball). L'espèce est connue en Amérique du Nord dans la plupart des États des États-Unis et l'est du Canada (Vidano, 1966 ; Maixner et al., 1993).
Elle a été identifiée pour la première fois en Europe en 1958 dans un vignoble de Gironde (Bonfils & Schvester, 1960).
Ensuite, elle s'est répandue vers le nord, le sud et l'est. Elle est actuellement présente dans la totalité des vignobles français à l'exception de ceux de Lorraine et d'Alsace ; dans toutes les provinces d'Italie, au nord du Portugal, en Tessin et Suisse romande ; en Slovénie, Croatie, Autriche, Hongrie et Serbie.
Ainsi, l'espèce s'est installée d'ouest en est dans une zone climatique ayant des hivers froids et des étés longs, entre la latitude nord de la Loire, de la Suisse, de l'Italie du Nord et de la Slovénie (environ 47 °N) et celle de la Corse, Nord Portugal et extrême sud de l'Italie (environ 42 °N).
On peut faire l'hypothèse que son cycle biologique s'est adapté à des étés plus courts et que son extension progressive vers le nord n'est pas achevée (Maixner & Boudon-Padieu, 2007). Au contraire, des hivers doux défavorables à la régularité des éclosions pourraient limiter la colonisation. L'introduction dans les vignobles européens d'oeufs de S. titanus, portés par les bois et plants commercialisés à de grandes distances, est la cause de cette vaste dispersion, comme cela a été démontré par deux études de typage génétique des insectes.
Distribution de la flavescence dorée
La flavescence dorée est largement présente en France dans toutes les régions viticoles du Sud et du Sud-Ouest. Plus de 300 000 ha de vignobles étaient placés en lutte obligatoire en 2000 (Roure, 2000). Des foyers se sont ensuite déclarés en Savoie en 2000 et au nord de la Provence en 2001, avec des incursions limitées à quelques ceps en Beaujolais (1996) et Bourgogne (2004).
À partir de 2011, de sévères attaques ont été à déplorer en Sud-Bourgogne, en région PACA, tandis qu'elles continuent leur progression en Charente et Aquitaine. En Italie du Nord, la flavescence dorée a été identifiée en Val d'Aoste, Piémont, Lombardie, Trentin-Haut-Adige, Vénétie, Frioule, Ligurie et Émilie-Romagne, ainsi qu'en Ombrie, Marches et Toscane.
Symptômes et étiologie de la flavescence dorée
Symptômes
La flavescence dorée et les autres jaunisses de la vigne produisent des symptômes semblables sur V. vinifera. Ils évoquent une mauvaise circulation de la sève et le transport défectueux des produits de la photosynthèse des feuilles vers les rameaux : enroulement des feuilles, anomalies de coloration des limbes et des nervures, absence partielle ou totale de stockage des réserves, manifestée par des sarments flexueux et le dépérissement d'une partie ou de la totalité de la souche (photos 1 et 2).
Lorsque la maladie se manifeste précocement, les fleurs se dessèchent et les fruits sont alors absents. Une manifestation plus tardive entraîne le flétrissement des baies à partir de la rafle et le défaut de maturité (photo 4 page suivante).
Étiologie
Les phytoplasmes sont des petites bactéries sans paroi (classe des mollicutes). Elles n'ont jamais été cultivées en milieu artificiel car ce sont des parasites obligatoires. Autrement dit, elles dépendent de l'insecte vecteur et de la plante-hôte pour leur multiplication. Leur étude nécessite donc des dispositifs expérimentaux très complexes.
Depuis leur découverte, en 1967, des phytoplasmes ont été associés à plusieurs dizaines de maladies des plantes. Leurs vecteurs ne sont pas tous connus.
Les jaunisses de la vigne ont été associées à plusieurs phytoplasmes différents dans plusieurs pays. Parmi eux, la flavescence dorée représente un cas où l'étiologie a été entièrement établie.
Bien que les phytoplasmes ne soient pas cultivables et que, par conséquent, le postulat de Koch n'ait pu être satisfait au sens littéral, le phytoplasme de la flavescence dorée a été observé et détecté de façon spécifique aussi bien dans ses vecteurs naturel et expérimental que dans les vignes malades, naturellement infectées ou inoculées expérimentalement à l'aide de vecteurs infectieux (Caudwell et al., 1971 ; Boudon-Padieu et al., 1989 ; Lherminier et al., 1990 ; Meignoz et al., 1992 ; Daire, 1994).
Interactions plante-phytoplasme
Les phytoplasmes sont des bactéries habitant exclusivement le phloème des plantes (photo 5).
Il a été montré sur des plantes inoculées expérimentalement par un insecte vecteur infectieux (la fève Vicia faba) ou par la greffe (la pervenche de Madagascar Catharanthus roseus), que les phytoplasmes descendent d'abord vers les radicelles, s'y multiplient puis remontent en envahissant le collet et l'ensemble du système racinaire ; ensuite, ils gagnent les jeunes organes en développement et se multiplient activement dans les nervures des jeunes feuilles (Kuske & Kirckpatrick, 1992 ; Lherminier et al., 1994).
Le phytoplasme de la flavescence dorée est présent dans les tissus de la vigne infectée selon une répartition irrégulière et à de faibles concentrations.
Les désordres cytologiques observés en microscopie électronique peuvent toucher la totalité d'un faisceau libérien dont les tubes criblés sont nécrosés. Mais ces désordres n'affectent souvent que quelques-uns des faisceaux d'un même rameau ou d'une même nervure. La plante semble réagir à l'infection, et affecter les phytoplasmes qui apparaissent coagulés et déformés (Meignoz et al., 1992 ; Credi, 1994a, 1994b).
D'autres tubes libériens ne contiennent que quelques cellules de phytoplasmes et conservent une apparence normale.
De ce fait, des sarments normalement aoûtés peuvent contenir des phytoplasmes latents.
Phytoplasmes latents, rémissions
Le prélèvement de tels sarments sur des vignes-mères contribuera à la diffusion de la flavescence dorée à distance par des boutures infectées (Caudwell & Larrue, 1979).
Toutes les variétés de V. vinifera sont sensibles à la flavescence dorée mais manifestent des degrés divers de sensibilité à la maladie.
Une rémission naturelle, ou rétablissement, peut être observée sur des ceps malades protégés de nouvelles inoculations. Ce rétablissement, décrit par Caudwell pour la flavescence dorée en 1961, se manifeste pour toutes les jaunisses de la vigne.
Une étude par Maixner (2011) dans le cas du bois noir montre que certaines plantes atteintes au vignoble se rétablissent de façon définitive après une période intermédiaire durant laquelle elles restent porteuses et susceptibles d'exprimer à nouveau des symptômes. Le taux de rétablissement dépend du cépage. Le rétablissement ne protège pas des réinoculations. Un cep rétabli inoculé peut à nouveau exprimer la maladie.
En ce qui concerne les porte-greffes, une étude décisive par Caudwell et al., (1994) a montré que des vignes-mères de diverses variétés étaient porteuses d'une infection latente et capables de transmettre par greffe la flavescence dorée à des greffons sensibles de V. vinifera. Les pieds-mères étudiés ont maintenu cette infectivité latente pendant plusieurs années.
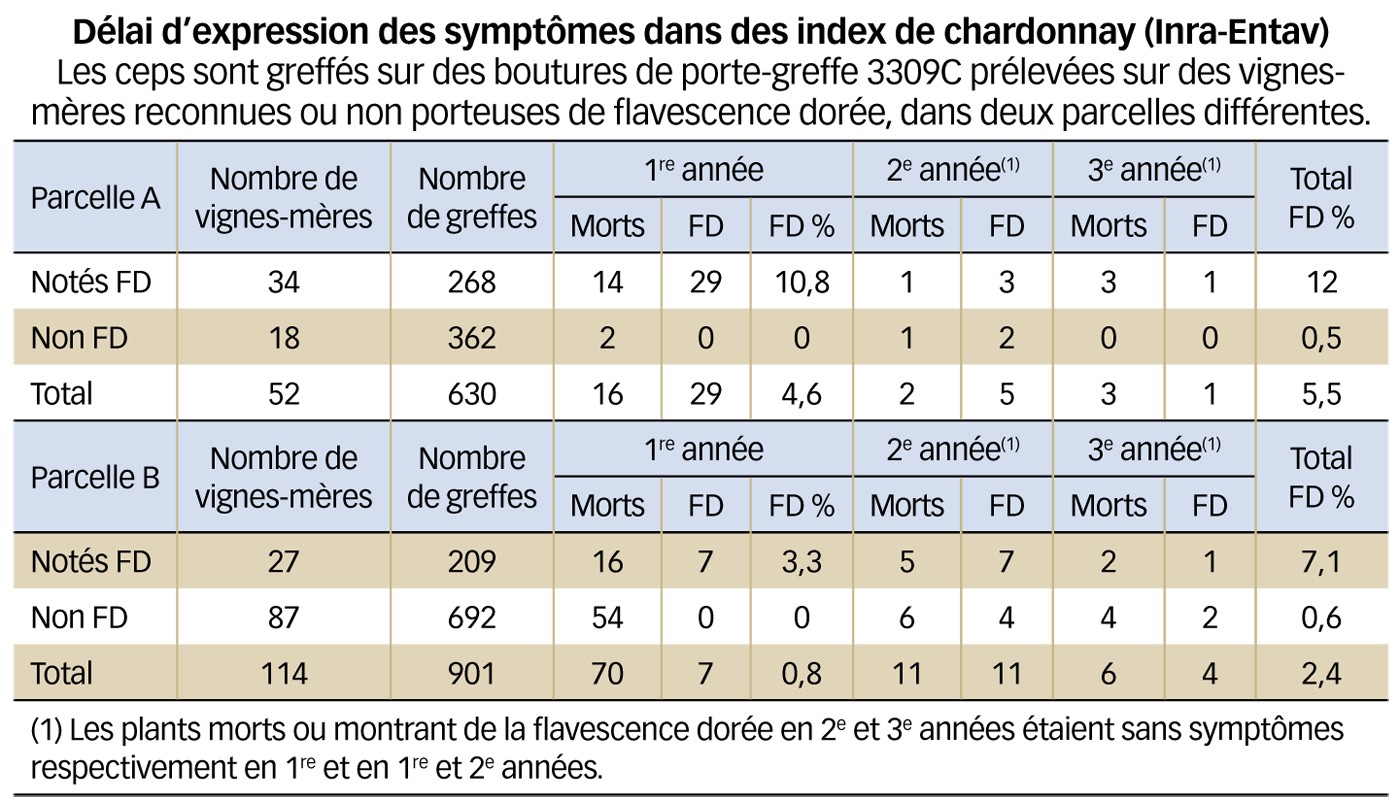
Le taux de transmission par des boutures de porte-greffe prélevées sur une même plante-mère, est imprévisible (Figure 1), allant de 6 % à 80 % selon l'état infectieux de la plante (Caudwell et al., 1994).
Sur la base de ces éléments, on peut postuler que le délai d'expression retardée après contamination, l'intensité des symptômes et le processus du rétablissement d'un cep malade dépendent non seulement du cépage du V. vinifera mais également de la variété du porte-greffe de l'assemblage.
Les boutures greffées sur le matériel porte-greffes infecté expriment le plus souvent leurs symptômes dès la pépinière.
Cependant, quelques greffés-soudés peuvent demeurer porteurs sains pendant plusieurs années. L'expérimentation a montré que l'expression de symptômes peut être retardée jusqu'à trois ans (voir tableau) ; en outre, des observations au vignoble suggèrent qu'une latence de cinq à sept ans pourrait advenir.
Imprévisible phytoplasme
En résumé, il semble que les réactions de défense des ceps soient irrégulières ; elles dépendent du cultivar ou du cépage des deux partenaires greffon et porte-greffe. Étant donné que ni l'état de colonisation par le phytoplasme ni sa répartition dans la plante ne sont prévisibles, une réponse négative à l'indexage biologique ou au diagnostic au laboratoire n'est pas fiable.
En outre, le titre en phytoplasmes dans les tissus de la plante porteuse peut demeurer quelque temps sous un « seuil d'expression de symptômes » et sans doute aussi sous le seuil de sensibilité des méthodes moléculaires de détection.
Biologie de la cicadelle vectrice et de la transmission par le vecteur
Un insecte très lié à la vigne
Une fois exprimés les premiers symptômes dans une parcelle de vigne, la diffusion de la flavescence dorée au sein de cette parcelle est extrêmement rapide. L'activité principalement ampélophage de son vecteur spécifique, la cicadelle S. titanus, et la grande efficacité de vection en sont la cause (Bressan et al., 2006b).
S. titanus est une espèce monovoltine (une seule génération par an) possédant cinq stades larvaires.
Les femelles gravides insèrent leurs oeufs dans les craquelures de l'écorce des bois de deux ans ou plus. Les éclosions commencent en France début mai et peuvent durer jusqu'à huit semaines. La durée moyenne de vie larvaire entre deux mues est de dix jours. Les larves aptères se nourrissent sur la face inférieure des feuilles.
En fonction des conditions climatiques, les premiers adultes peuvent apparaître dès la mi-juillet ; cependant, des larves de cinquième stade peuvent être observées jusqu'à la fin août. Les adultes femelles apparaissent et persistent plus tard que les mâles et déposent leurs pontes jusqu'à leur mort en septembre. Les oeufs passent l'hiver sous l'écorce des bois de deux ans ou plus.
Cette cicadelle est un vecteur, hélas, très efficace
La propagation du phytoplasme de la flavescence dorée a été étudiée dans le corps d'un vecteur expérimental (Lefol et al., 1994). Les phytoplasmes acquis pendant la prise de nourriture pénètrent dans le tube digestif avec le bol alimentaire.
Après l'acquisition, une période de latence précède l'état infectieux de l'insecte. Cette latence est le temps nécessaire aux phytoplasmes, présents dans l'intestin, pour envahir la cavité générale, traverser les parois cellulaires et pénétrer dans divers organes, dont les glandes salivaires. Dès lors, l'insecte devient infectieux. Cette généralisation dans l'organisme est accompagnée de la multiplication des phytoplasmes, en particulier dans les cellules sécrétrices de salive (Lherminier et al., 1990 ; Lefol et al., 1994).
Comme le phytoplasme n'est pas transmis par l'adulte à sa descendance (transmission verticale), la diffusion d'un phytoplasme n'est possible que si chaque génération de l'espèce vectrice est confrontée à une acquisition.
Ce schéma de propagation du phytoplasme de la flavescence dorée dans le corps du vecteur a été vérifié dans des spécimens de son vecteur naturel S. titanus (Boudon-Padieu et al., 1989). La latence mesurée est de quatre semaines environ. C'est en combinant la date des premières éclosions et ce délai de latence qu'est déterminée la date des premiers traitements insecticides dans la lutte indirecte contre la flavescence dorée.
Cette transmission sur le mode « persistant, circulant, multipliant » suggère la présence de facteurs de reconnaissance spécifique entre les cellules du phytoplasme et celles de l'insecte vecteur.
Ces facteurs seraient nécessaires au passage des phytoplasmes à travers la paroi intestinale et à leur pénétration dans les cellules sécrétrices des glandes salivaires (Lefol et al., 1994 ; Bressan et al., 2006a).
Toutefois, la présence du phytoplasme de la flavescence dorée dans le corps des insectes S. titanus n'est pas anodine.
Il a été démontré que les spécimens infectés, mâles et femelles, ont une longévité réduite par rapport à leurs comparses non infectés ; en outre, les femelles infectées sont moins prolifiques que celles non infectées (Bressan et al., 2005).
Ces derniers éléments suggèrent que l'association entre le phytoplasme de la flavescence dorée et son vecteur est « récente » au regard du calendrier de l'évolution.
Biologie de la flavescence dorée
Les Vitis sp. sont aussi bien les plantes-hôtes du vecteur que du phytoplasme de la flavescence dorée, caractères combinés qui favorisent un cycle d'infestation rapide du vignoble (Bressan et al., 2006b).
L'acquisition et l'inoculation par le vecteur se produisent en été ; l'incubation se déroule dans les ceps inoculés pendant l'automne, l'hiver et le printemps (Figure 2).
Le taux annuel de multiplication du nombre de ceps exhibant des symptômes au sein d'une parcelle est très élevé. Par exemple, le nombre de ceps affectés dans une parcelle de Savoie a été multiplié par vingt entre 2000 et 2001 (SDQPV Rhône-Alpes, comm. pers).
La période des acquisitions commence dès le début des éclosions. Une larve de premier stade née sur une souche infectée peut acquérir le phytoplasme de la flavescence dorée dès son premier repas.
Les spécimens infectés deviennent infectieux quatre à cinq semaines après cette première acquisition (durée de la latence) quel que soit le stade de développement atteint entre temps, et ils conservent leur pouvoir infectieux jusqu'à leur mort.
Ainsi, la période des transmissions débute un mois après la naissance du premier spécimen (soit début juin) et dure jusqu'à la mort des adultes en automne. Acquisition et transmission sont des événements de très haute efficacité (Bressan et al., 2006).
Les ceps de vigne infectés par des phytoplasmes constituent des réservoirs disponibles pour l'acquisition par l'insecte vecteur pendant sa période d'activité, puis des réservoirs de conservation des phytoplasmes durant le stage oeuf hivernant de l'insecte.
Comme les symptômes n'apparaîtront sur les nouvelles souches inoculées que pendant l'été de l'année suivante, le nombre de plantes montrant des symptômes à un moment donné pendant la phase d'expansion épidémique est une sous-estimation du nombre total de plantes infectées prenant en compte les plantes « en incubation ».
Les jeunes feuilles qui se développent au printemps sur les souches inoculées pendant l'été précédant contiennent des phytoplasmes accessibles par prise de nourriture par les larves, avant même que les symptômes ne soient visibles sur ces feuilles.
En outre, les vignes-mères infectées portent des bois en incubation qui fourniront des greffons porteurs (Figure 2) (Caudwell et al., 1994).
Conclusion
Les caractères les plus frappants des épidémies de flavescence dorée résultent de la spécificité et de la biologie de sa cicadelle vectrice.
Cette espèce n'est qu'une des nombreuses espèces américaines appartenant au même genre et ayant une éthologie comparable, vivant sur des Vitis sauvages (V. riparia) (Nusillard & Malausa, comm. pers.). Il est remarquable que S. titanus ait trouvé en Europe des conditions des plus favorables pour sa multiplication et son extension sur V. vinifera, et soit devenu une des plus importantes cicadelles présentes sur la vigne dans nos pays.
Les caractères biologiques décrits ici soulèvent de nombreuses questions sur l'origine et les conditions d'apparition de la flavescence dorée.
La lutte - nous l'exposerons dans le prochain article - peut être directe ou indirecte selon que l'on tente d'éradiquer les plantes malades ou le vecteur. Mais elle doit, dans tous les cas, s'appuyer sur une prophylaxie éclairée.
C'est là un point extrêmement controversé. Notre message aux praticiens a toujours été de réduire par tous les moyens les populations de vecteur avant même l'arrivée de la maladie. Mais il a rarement été entendu, car le vecteur, très discret, ne cause pas de dégâts directs.
En outre, les praticiens ont souvent tendance à sous-estimer la gravité de la flavescence dorée tant qu'ils ne sont pas confrontés à sa réalité.
Fig. 1 : Impossible de prévoir les boutures contaminées
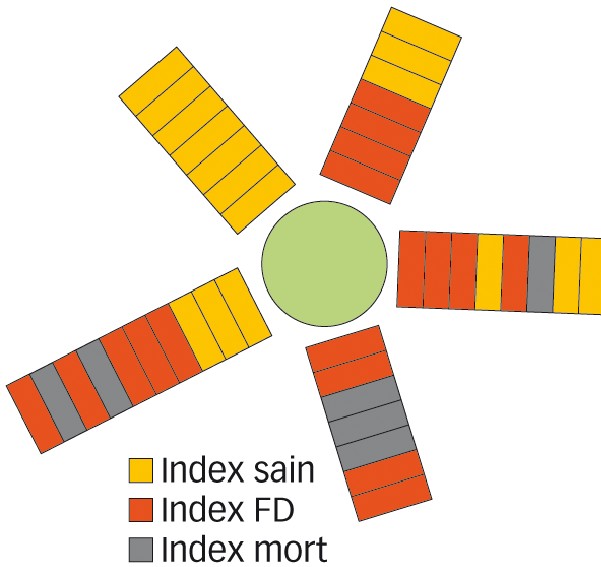
Analyse par indexage de la distribution du phytoplasme de la flavescence dorée dans les sarments d'une vigne-mère de porte-greffe. Les boutures ordonnées de porte-greffe sont indexées avec des greffons de chardonnay sensible à la maladie. On voit l'irrégularité de sa répartition...
(collaboration Entav-Inra).
Fig. 2 : La lenteur des symptômes à apparaître... accélère l'épidémie
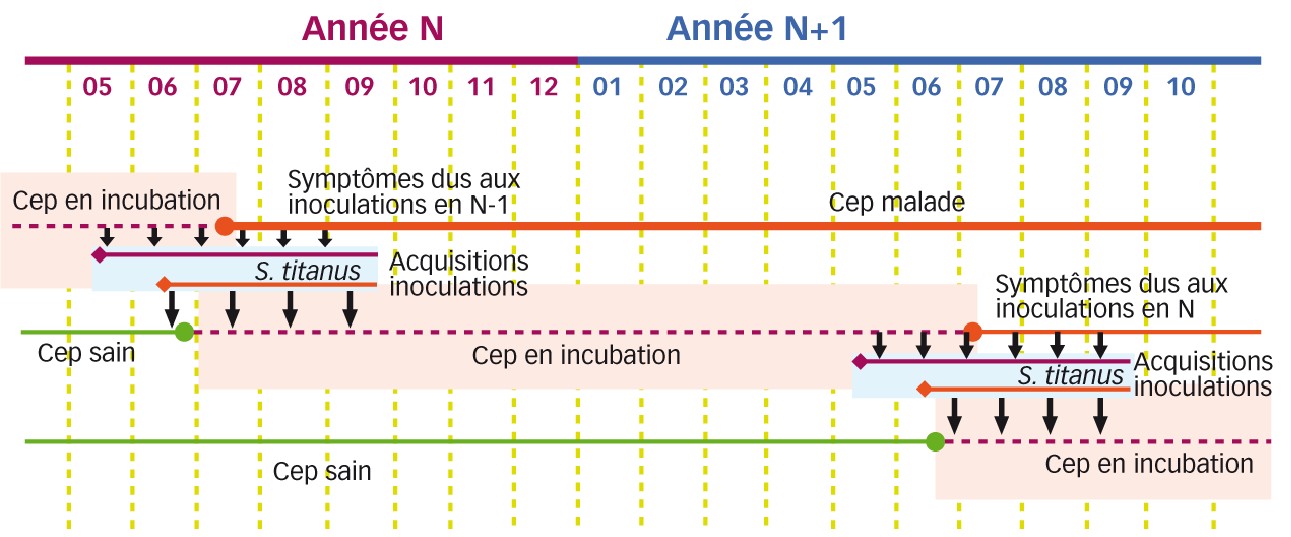
Biologie de la transmission de la flavescence dorée par S. titanus et incubation hivernale du phytoplasme dans les ceps. Les ceps en incubation sont des « caches » de phytoplasme.

