Imprimer
Imprimer
Dégâts sur ail (ph.F. Villeneuve, Ctifl

Trois phéromones de taupins disponibles ; testées dans le même type de piège, toutes attirent A. lineatus et A. sordidus, mais avec des niveaux de capture différents.
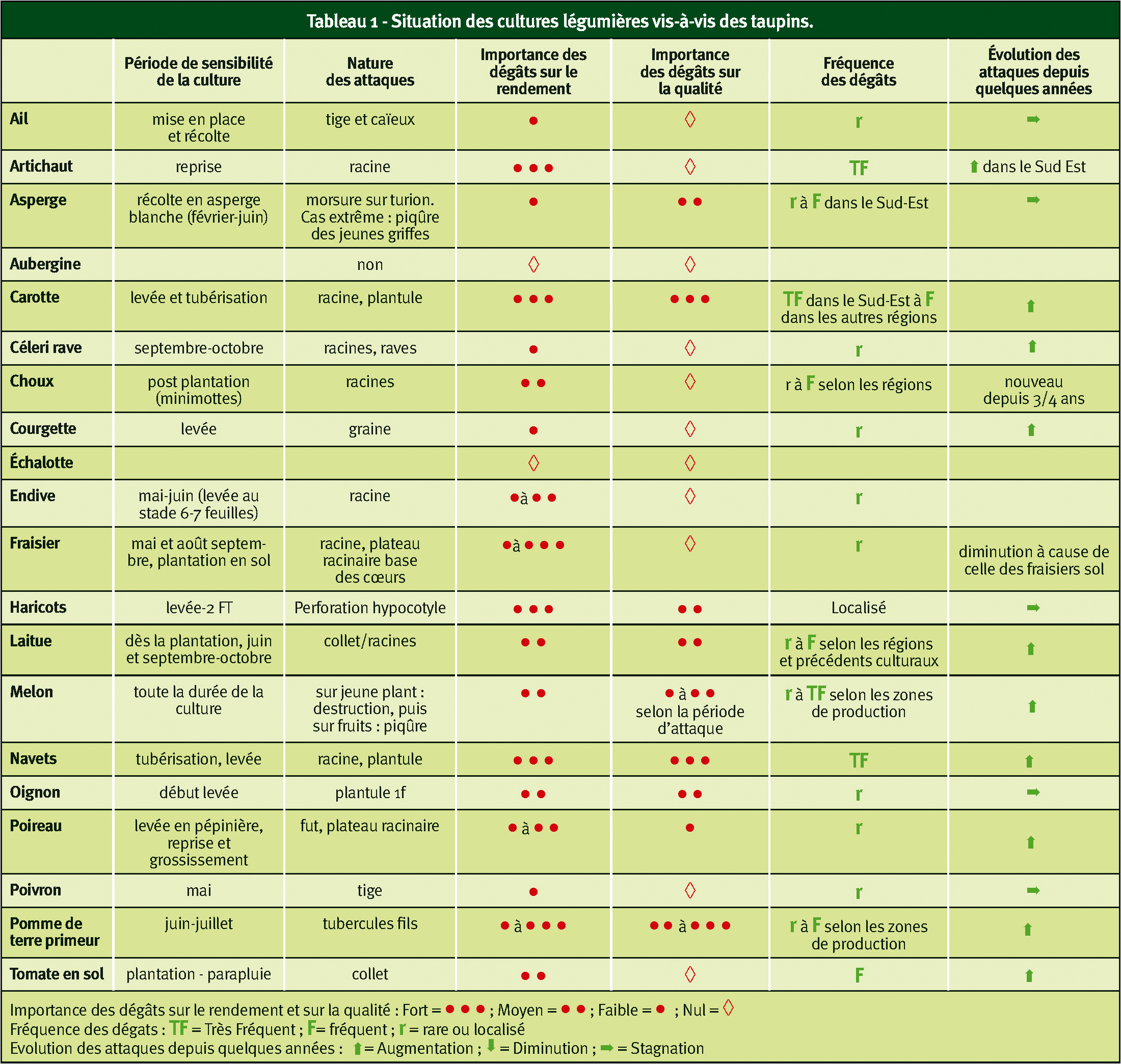
, en médaillon page précédente sur ail. Le tableau ci-dessous montre l'importance du problème en cultures légumières. ph. P. Mention, Ctifl")
Dégâts de larves de taupin : ci-dessus sur melon (même si on ne les voit pas souvent en surface, les larves ne s'attaquent pas qu'aux organes enterrés des plantes cultivées), en médaillon page précédente sur ail. Le tableau ci-dessous montre l'importance du problème en cultures légumières. ph. P. Mention, Ctifl
 mais pas au même niveau. ph. F. Villeneuve, Ctifl")
À gauche, le piège Yatlor f dérivé du piège de type Rebell mis au point par le Dr Lorenzo Furlan. à droite, le piège de type « pit fall ». Tous deux ont été comparés à Lanxade. Ils ont piégé en majorité des A. sordidus, ce qui est normal pour la région, avec une dynamique comparable (pics de capture aux mêmes périodes) mais pas au même niveau. ph. F. Villeneuve, Ctifl
La recrudescence des dégâts de taupins touche les grandes cultures, mais aussi les cultures légumières. Aussi le Ctifl, Institut technique des fruits et légumes, se penche sur la question en particulier en testant le piégeage des adultes de taupins sur son site de Lanxade, en Dordogne, depuis 2004. Certains résultats de ce travail sont surprenants : ainsi, le taupin vole plus loin qu'on ne croit. D'autres semblent décourageants mais sont utiles : non-relation entre piégeage des adultes et pression des larves dans le sol, médiocres résultats de certaines méthodes de lutte : on sait maintenant comment ne pas les appliquer, reste à les tester autrement. Enfin il reste un espoir (à confirmer !) pour le piégeage de masse.
Actuellement, les dégâts de taupins augmentent sur de nombreuses cultures suite à des changements de pratiques culturales : travail simplifié du sol, jachères, insecticides retirés et traitements en plein remplacés par des traitements localisés ou de semences... On note aussi le développement d'une espèce à cycle court, Agriotes sordidus. Les dégâts touchent même des parcelles et cultures qui semblaient indemnes ou peu sensibles, notamment en cultures légumières (Tableau 1).
Les phéromones changent la donne
Les phéromones, outil intéressant pour surveiller les populations
En parallèle, la découverte de phéromones (Borg-Karlson et al., 1988 ; Yatsynin et al., 1996 ; Tóth et al., 2003) a modifié les perspectives en termes de protection comme de prévision des risques et d'amélioration des connaissances de la biologie sur ces ravageurs.
En effet, chez les taupins, seuls les adultes sortent du sol. Et leur observation directe n'est pas aisée du fait de leur mode de vie : comportements crépusculaire et nocturne, vols assez peu fréquents, relative brièveté de la vie imaginale par rapport à la vie larvaire... Une solution pour les repérer est donc de les piéger.
Autrefois, le piégeage utilisait des pièges passifs, notamment les bottillons (Aguilar, 1948). Les études avaient abouti à estimer le vol des taupins comme une activité exceptionnelle avec déplacements de faibles amplitudes (Aguilar, 1961 ; Wilkinson, 1963). Aujourd'hui, les phéromones rendent les pièges attractifs donc plus efficaces.
Les conditions et possibilités d'utilisation de ces phéromones obligent à mieux connaître certains éléments, tels que la spécificité, le sexratio... (Villeneuve et al., 2006 et 2008). Des travaux similaires ont été réalisés dans de nombreux pays. Le niveau de spécificité dépend de l'espèce d'Agriotes considérée, mais aussi de la quantité de taupins capturés (Vernon et Tóth, 2007 ; Blackshaw et al., 2009...). Des travaux au Royaume Uni ont même abouti à une tentative de grille d'évaluation du risque pour les cultures de pomme de terre (Anonyme, 2006).
Plusieurs phéromones et des pièges variés
Aujourd'hui, les phéromones disponibles sur le marché viennent de sources différentes et les pièges dédiés à la capture des adultes de taupins sont plus ou moins spécifiques.
La comparaison entre les sources de phéromones et les types de pièges montre des différences importantes, portant généralement plus sur les niveaux que sur les dynamiques de piégeage (Villeneuve et al., 2008 ; Villeneuve et Latour, 2009). Villeneuve et al. (2008) ont décelé une interaction entre composés phéromonaux lorsque l'on veut piéger des espèces différentes dans une même parcelle.
La découverte des phéromones et la mise au point de pièges ont permis des avancées et confirmé la difficulté pour un non-expert d'identifier les espèces présentes dans un sol.
Par ailleurs, les méthodes d'estimation des risques à partir d'une évaluation des populations de larves sont longues, fastidieuses et coûteuses en main-d'œuvre avec des seuils de risques très bas pour les cultures. L'utilisation des pièges à phéromones semble pouvoir remédier en partie à ces inconvénients car la détection des populations et l'effort d'échantillonnage sont inférieurs. Mais il faut que ces données ne fassent pas illusion par une apparente précision des niveaux de captures.
L'objectif de notre travail est de mieux comprendre certains facteurs de variations du piégeage des adultes de taupins et d'évaluer certaines pistes de protection des cultures utilisant des pièges à phéromones.
Sept ans sur un site
Divers essais ont été menés entre 2004 et 2010 sur le site Ctiflde Lanxade (près de Bergerac, 24), dans plusieurs parcelles avec une distance moyenne entre elles de 200 à 500 m.
Deux types de phéromones ont été utilisés : un d'origine grecque (Novagrica-Entomos) qu'on ne change pas en cours de saison et l'autre d'origine hongroise (Csalomon®, Plant Protection Institut of Hungarian Academy of Science, aimablement fourni par Syngenta Agro) qu'il faut changer tous les 45 jours. La charge en phéromones des capsules et la nature exacte du contenant ne sont pas connues.
Deux types de pièges ont été testés : un piège adapté aux taupins par Furlan dit « Yatlor f » dérivé du piège de type Rebell et un piège de type « Pitfall » mis au point pour le piégeage du charançon du bananier.
Selon les essais, les relevés des pièges ont été effectués à la cadence d'une ou deux fois par semaine. Les adultes capturés sont placés dans une solution d'alcool à 75° avant d'être comptabilisés, identifiés et sexés. Les identifications ont été réalisées par examen des génitalias en se référant à Leseigneur (1972).
Le suivi des conditions climatiques a été réalisé grâce à une station météorologique Pulsonic.
Diversité constatée
Espèces présentes sur le site de Lanxade
Le suivi depuis 2004 des captures d'Agriotes sur le site de Lanxade dans une parcelle identique de 2 600 m² a permis d'apporter quelques précisions sur leur biologie (Figure 1).
L'espèce la plus présente est A. sordidus. Sa période de capture, beaucoup plus longue que celle des autres espèces présentes sur le site, va de fin mars aux premières gelées, même si à ce moment-là les captures sont très faibles.
La deuxième espèce est A. lineatus avec une émergence légèrement plus tardive que celle d'A. sordidus et une fin de vol fin juillet.
Pour sa part, A. sputator est bien plus discrète, ses émergences légèrement plus tardives qu'A. lineatus et sa fin d'activité identique.
Quelques A. obscurus sont présents (4 individus en 2010, et 2 en 2007) ainsi que quelques individus A. gallicus – non considéré comme ravageur des cultures – rencontrés dans les pièges avec phéromone pour A. sordidus.
Variations observées : rôle de la température
On observe une forte variabilité des captures d'imagos d'A. sordidus sur une même parcelle d'année en année (Figure 2). Le même type de fluctuation est observé pour A. lineatus (résultats non présentés).
On peut imaginer que ces fluctuations sont liées à des conditions spécifiques de l'année qui influent soit sur l'émergence des imagos de leur logette, soit sur l'activité de recherche d'un partenaire, ou à des variations de niveaux de populations des larves suite à des événements divers (forte mortalité des œufs ou des jeunes larves, conditions climatiques adverses...). Ce résultat va contre l'idée couramment répandue que les cohortes annuelles de taupins sont de tailles similaires.
Certains facteurs climatiques intervenant sur l'activité de vol ont déjà été mis en évidence comme la température, l'humidité relative et l'ensoleillement (Aguilar, 1961).
L'analyse des conditions climatiques enregistrées sous abris de 2004 à 2010 et des niveaux de captures hebdomadaires a permis de rechercher les facteurs climatiques pouvant intervenir. Ainsi, nous n'avons pas pu mettre en évidence d'influence du vent et relativement peu des précipitations qui ne sont qu'un indicateur indirect de l'humidité du sol. En revanche, nous avons pu observer une influence de la température (Figure 3).
Il n'y a pas d'activité de vol d'A. sordidus en dessous de 10 °C. Ceci semble concorder avec les observations de Furlan (2004) indiquant que les activités des larves s'arrêtent en dessous de 9 °C. Entre 10 ° et 14 °C, l'activité de vol est relativement faible. Mais cette activité augmente très nettement à partir de 14 °C pour rester soutenue quelle que soit la température.
Concernant A. lineatus, les mêmes observations peuvent être faites : pas de piégeage en dessous de 10 °C, faible niveau de captures entre 10 et 14 °C, activité nettement augmentée à partir de 14 °C (résultats non présentés).
Différences entre parcelles
Des fortes variations de niveaux de captures s'observent entre des parcelles proches.
Par exemple, le suivi des captures d'A. sordidus et A. lineatus sur deux parcelles distantes de 200 m en 2009 pendant 12 semaines consécutives, nous montre une nette différence entre les niveaux de captures (A. sordidus parcelle n°1 : 642 individus, parcelle n°2 : 109 individus ; A. lineatus parcelle n°1 : 163 individus, parcelle n°2 : 58 individus).
De même, la dynamique des captures diffère (Figure 4).
Les adultes ne disent rien des larves
Il est légitime de se demander si ces différences de captures des adultes se traduisent par des différences du nombre de larves présentes dans le sol. L'estimation du nombre de larves a été effectuée avec le piège Acta.
Nous n'observons pas de relation entre les niveaux de captures des adultes et les populations de larves : sur la parcelle n°1, ayant le niveau le plus élevé de captures d'imagos, nous avons 0,5 larve par piège (moyenne de 40 pièges) et sur la parcelle n°2, 0,4 larve par piège. Deuxième point, la majorité des larves capturées appartiennent à l'espèce A. lineatus (97 % des larves), alors que l'espèce d'adultes la plus présente dans les pièges est A. sordidus.
Ces observations sont corroborées par des résultats comparables d'autres équipes de recherche (Blackshaw et Vernon, 2006 ; Blackshaw et al., 2009). Elles montrent combien, dans une parcelle, sont complexes les relations entre les taupins mâles – dont la quantité est estimée au travers des pièges à phéromones – et les larves de taupins présentes.
Certains points de la biologie des taupins ne sont pas clairement établis, par exemple la distance qu'ils peuvent parcourir. Existe-t-il des différences entre espèces ? Réagissent-elles de la même façon à la même quantité de composés phéromonaux ? Ont-elles les mêmes températures optimum pour le déplacement ?
Noronha et al. (2008) indiquent que la corrélation entre l'abondance relative et la distribution spatiale des larves et des adultes dépend de l'espèce d'Agriotes considérée.
Mobilité insoupçonnée
L'utilisation des pièges à phéromones nous a montré que les déplacements des mâles d'Agriotes sp. sont beaucoup plus fréquents et importants que ne le laissaient supposer les travaux antérieurs.
De plus, dans une étude de recapture d'individus marqués, nous enregistrons, pour les mâles d'A. sordidus lâchés à 20 m des pièges, un taux de recapture de 25 %, et surtout un taux de 17 % de recapture 23 jours après le lâcher à une distance de 40 m. Nous avons même recapturé un individu à plus de 800 m de son lieu de lâcher.
Hicks et Blackshaw (2008) ont montré des différences entre espèces d'Agriotes : A. lineatus se déplace plus qu'A. obscurus qui elle-même se déplace plus qu'A. sputator.
Perspectives de protection
Insecticides visant les adultes ?
La découverte et la mise au point des pièges à phéromones pour les taupins ont ouvert des perspectives en terme de nouvelles méthodes de protection. Ainsi la connaissance des périodes et des intensités de capture, donc d'activité, permet d'envisager un changement de stratégie de protection chimique : celle-ci pourrait viser les adultes et non plus les larves.
Mais la méthode, testée aux Pays-Bas (Ester et al., 2002) et en France, a des limites. D'abord, l'efficacité ne peut pas être immédiate ; vu la biologie des taupins (cycles de 2 à 5 ans), seule une fraction de population potentielle est touchée, sans effet immédiat.
Ensuite, cela oblige à traiter des cultures n'ayant pas forcément besoin de protection. Enfin il faut disposer de produits efficaces contre les adultes. Les premiers résultats n'ont pas été concluants pour le moment.
Piégeage de masse ?
Compte tenu des niveaux de piégeage et de la mobilité moyenne des adultes, il est possible d'imaginer le piégeage de masse, qui a fait ses preuves sur d'autres ravageurs. L'effet ne peut pas être immédiat compte tenu des cycles biologiques. Cette protection doit s'envisager sur l'ensemble des cultures de rotation.
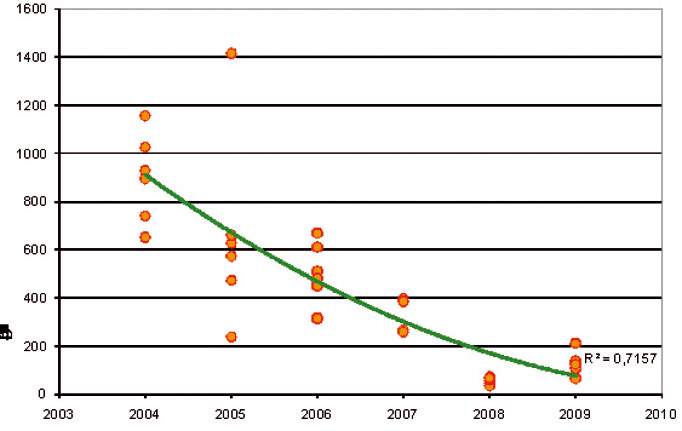
Depuis 2004, un piégeage est effectué sur la même parcelle (6 pièges pour 2 600 m²) sur le site de Lanxade. On y observe une réduction importante du nombre d'adultes d'A. sordidus capturés (Figure 5). Nous enregistrons une baisse régulière des niveaux de captures de 2004 à 2008. Mais ensuite le niveau semble s'être stabilisé, voire progresser légèrement : cela mérite d'être suivi.
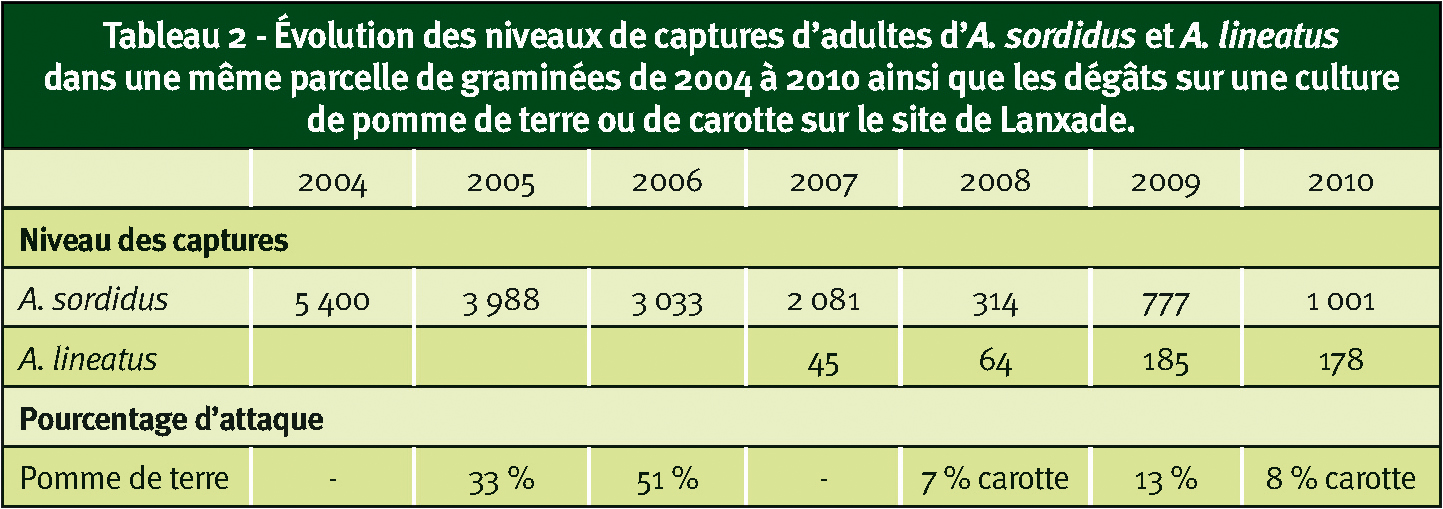
Concernant A. lineatus, également présent sur le site, 3 pièges ont été mis en œuvre depuis 2007 mais il n'y a pas de réelle évolution des individus piégés (Tableau 2).
Pour connaître l'impact de ce piégeage de masse, une culture « révélatrice » est mise en place chaque année, soit de pomme de terre, soit de carotte. Le niveau des dégâts observés sur pomme de terre est fortement réduit (près de 70 %) mais encore inacceptable pour un producteur. Sur carotte, il n'y a pas de réelle évolution des dégâts entre 2008 et 2010 (Tableau 2).
Phéromone et nématode auxiliaire associés ?
Enfin, on peut envisager le piégeage comme « attracticide », soit en contaminant la population de taupins par un agent biologique (nématode entomopathogène), soit en associant insecticide et phéromone (système attract and kill). La technique a été testée avec plus ou moins de succès sur divers ravageurs : charançon du bananier Cosmopolites sordidus, mouche de l'olive Bactrocera oleae, noctuelle de la tomate Helicoverpa armigera, carpocapse du pommier Cydia pomonella...
Les tentatives réalisées avec une souche d'Heterorhabditis megidis et une autre d'H. bacteriophora n'ont pas été concluantes. Nous n'avons pas pu mettre en évidence de contamination des adultes de taupin avec l'une ou l'autre des souches de nématodes entomopathogènes.
Rien d'opérationnel, mais des pistes à explorer
Ce qu'on a appris
La mise en œuvre du piégeage à phéromones des Agriotes sur le site de Lanxade a montré la présence des quatre espèces préjudiciables aux cultures, à des niveaux très différents : A. sordidus est la plus souvent piégée. Au contraire des observations antérieures, les mâles ont une activité de vol non négligeable, même si toutes les captures en pièges à phéromone ne surviennent pas après un épisode de vol.
Le piégeage commence à partir d'une température moyenne de 10 °C, mais reste faible jusqu'à 14 °C. Ce résultat contredit celui de Crozier (2007) indiquant une température minimum de 25 °C pour déclencher le vol chez A. obscurus et A. lineatus en conditions contrôlées.
Nous constatons de fortes variations interannuelles dans les niveaux de captures, mettant à mal l'idée de cohortes annuelles identiques pour une espèce en un lieu donné. Vu la durée de vie larvaire, de nombreux facteurs peuvent influencer les niveaux de populations.
Les niveaux de captures entre parcelles proches peuvent varier grandement. Ceci suggère qu'il existe des parcelles à risques plus ou moins forts et que le piégeage des adultes serait un moyen d'appréhender ce risque.
De plus, Blackshaw et Vernon (2006) ont montré une stabilité temporelle dans la structuration spatiale d'A. lineatus et A. obscurus en lien principalement avec les éléments stables du paysage.
Cela sous-tend qu'on pourrait établir une relation entre les niveaux de piégeage des adultes et de populations larvaires sur une parcelle donnée, et aussi que la répartition spatiale des mâles est représentative de celle des femelles. Enfin, l'identification des facteurs environnementaux favorables ou non aux taupins peut être une voie à explorer afin de manipuler leurs populations d'où leur impact sur les cultures.
Questions en suspens
Dans notre cas, il n'a pas été possible de voir de relation simple entre le niveau de captures des mâles et le niveau de population larvaire. Sur le site de Lanxade, l'espèce la plus capturée au stade adulte est A. sordidus, mais la plus capturée au niveau larvaire est A. lineatus : de 2007 à 2010, 680 adultes d'A. sordidus piégés (par piège, moyenne de 6 pièges) contre 177 A. lineatus, donc A. sordidus représentant 79 % de l'effectif. Or, au niveau larvaire, plus de 90 % des larves appartenaient à l'espèce A. lineatus. à ce stade de nos travaux, nous ignorons pourquoi : attractivité des pièges à phéromones à longue distance qui ferait venir des adultes depuis l'extérieur de la parcelle, réponse spécifique de chaque espèce au piège, inadaptation du piège ACTA pour capturer les larves ?
Les résultats de Hicks et Blackshaw (2008) montrent que l'attractivité des pièges à phéromones varie en fonction de l'espèce d'Agriotes considérée ; ceci peut jouer sur le nombre de pièges à mettre en œuvre par unité de surface.
Par ailleurs, l'activité de vol peut être sous-estimée. Les travaux de Hicks et Blackshaw (2008) ont un niveau de recaptures pour A. obscurus assez faible à partir de 32 m, pouvant indiquer la distance de vol de cette espèce. A contrario, les travaux Schallhart de et al. (2009), en utilisant des individus marqués au d13C, ont montré que l'espèce est capable de migrer sur 80 m. Enfin, en termes de protection, les premières tentatives d'utilisation des pièges à phéromones se sont révélées décevantes.
<p>* Ctifl, Centre de Lanxade, 28, route des Nébouts, 24130 Prigonrieux, villeneuve@ctifl.fr</p>
Figure 1 - Comparaison de l'activité moyenne de vols au cours des années 2004 à 2010 pour A. sordidus, A. lineatus et A. sputator sur le site de Lanxade (24).

Figure 2 - Fluctuations interannuelles et au cours de l'année de la dynamique de capture d' A. sordidus sur la même parcelle de 2005 à 2010 (moyenne de 6 pièges).
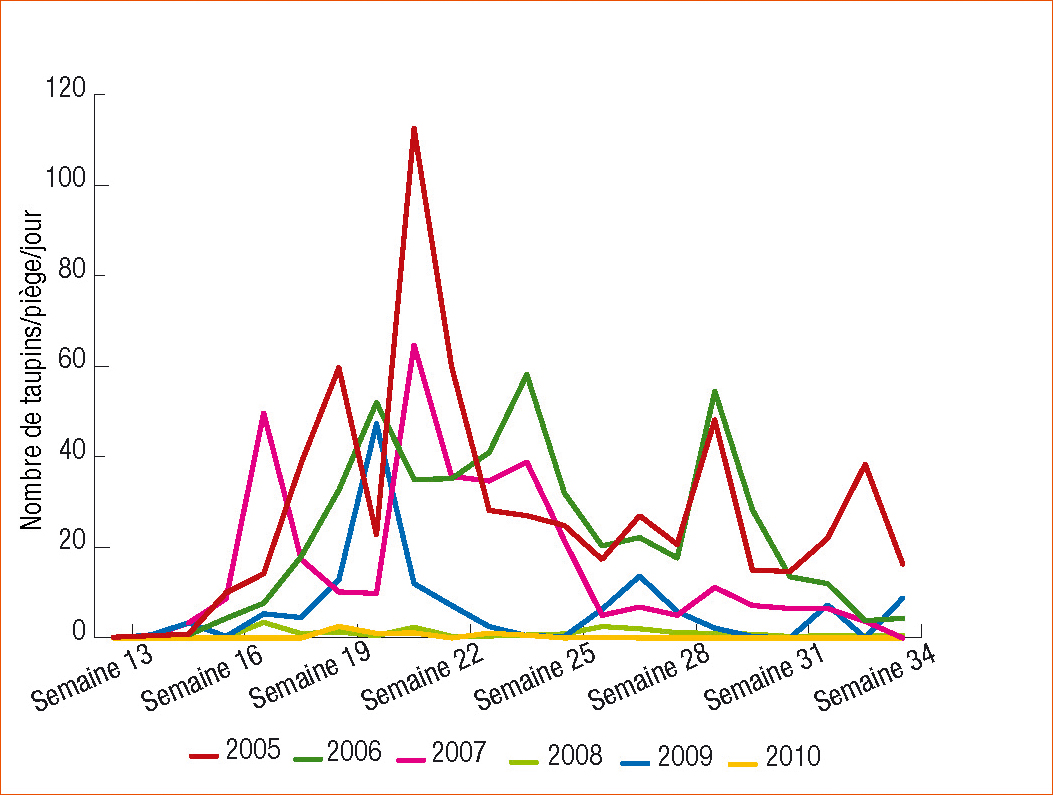
Figure 3 - Effets du vent (a) (en km de vent pour la semaine correspondante de captures), des précipitations (b) (en mm de pluie tombée dans la semaine de captures) et des températures (c) sur les niveaux de captures d'A. sordidus. Données de 6 pièges de 2004 à 2010 dans une même parcelle.
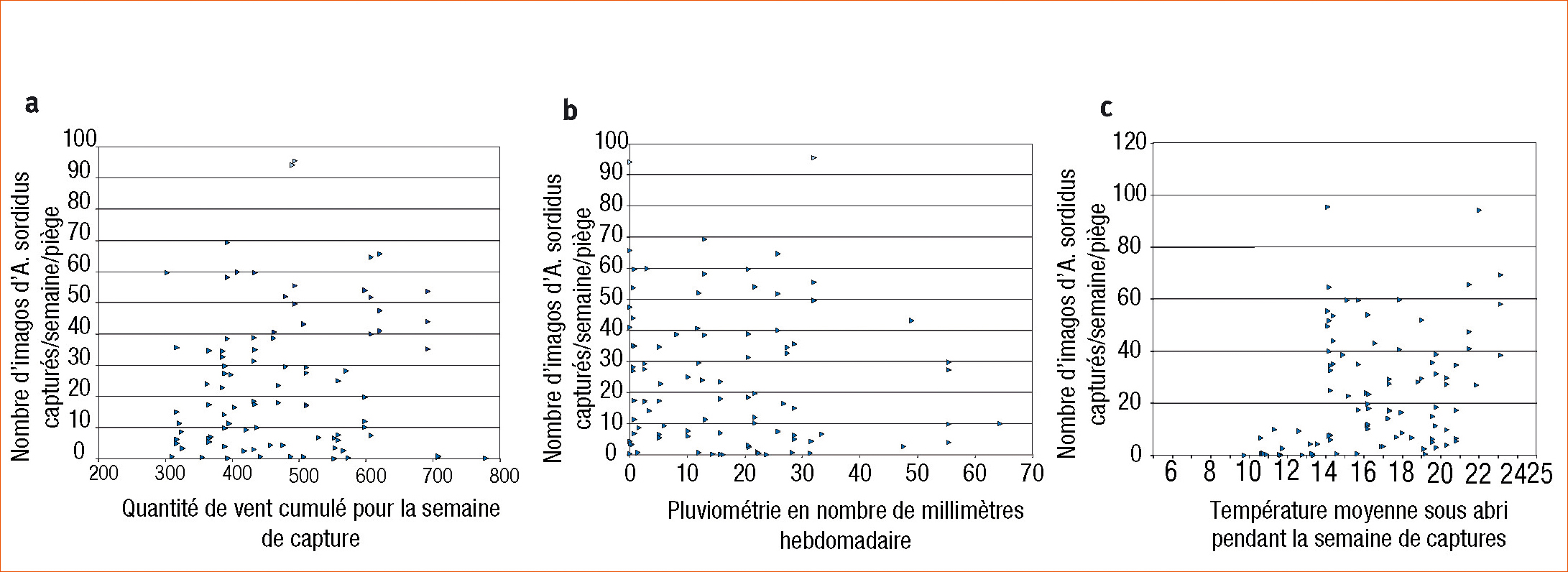
Figure 4 - Dynamique (et niveau) de captures des imagos d'A. sordidus et A. lineatus sur deux parcelles distantes de 200 m durant une période de 12 semaines en 2009.
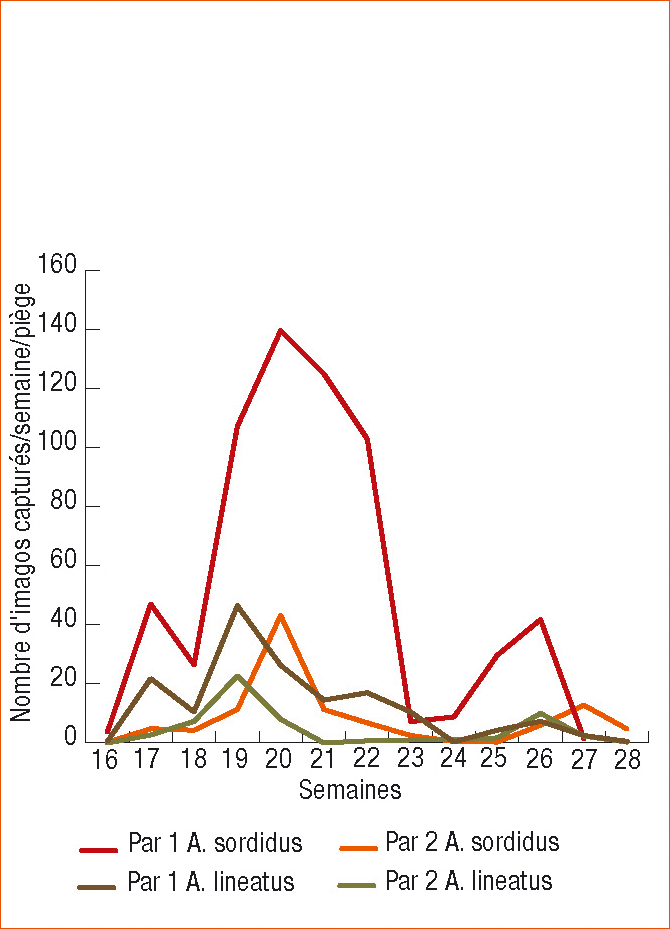
Figure 5 - Évolution des niveaux de captures d'adultes d'A. sordidus à l'aide de 6 pièges (essai de piégeage de masse) dans une parcelle de graminées de 2 600 m² de 2004 à 2009.