
 Imprimer
Imprimer")
ph. T. Verfaille (INHP Angers)
 et larves sur feuilles de platane. À droite (et en médaillon page précédente), décolorations occasionnées. Les nuisances sont ces dégâts directs, mais surtout l'émission de miellat et l'envahissement des maisons dont se plaignent les riverains. En bas à droite, l'avenue Jeanne-d'Arc à Angers, site d'une des expérimentations rapportées ici. Photos : T. Verfaille (INHP Angers)")
Les tigres en été : adultes (clairs et translucides) et larves sur feuilles de platane. À droite (et en médaillon page précédente), décolorations occasionnées. Les nuisances sont ces dégâts directs, mais surtout l'émission de miellat et l'envahissement des maisons dont se plaignent les riverains. En bas à droite, l'avenue Jeanne-d'Arc à Angers, site d'une des expérimentations rapportées ici. Photos : T. Verfaille (INHP Angers)
Face aux attentes du public, les professionnels de l'entretien des végétaux en ville cherchent à développer la protection biologique (Gutleben & Provendier, 2009). Ainsi, contre le tigre du platane Corythucha ciliata, on envisage d'utiliser des organismes entomopathogènes et des prédateurs. Le nématode Steinernema et la chrysope Chrysoperla lucasina ont été testés, d'abord séparément afin d'identifier les conditions de leur efficacité, puis en 2011 en stratégie combinée. Cet article décrit les travaux réalisés et leurs résultats. Et il montre qu'en lutte biologique aussi, les bonnes pratiques (ici, d'apport d'auxiliaires) c'est important !
Le tigre du platane, Corythucha ciliata (Say) (Hemiptera : Tingidae), a été identifié comme l'un des principaux problèmes phytosanitaires pour les gestionnaires d'espaces verts au sein des collectivités territoriales (Brunel, 2006 ; Chapin et al., 2006).
Pourquoi faut-il chasser le tigre ?
Quasi omniprésent
Originaire d'Amérique du Nord, il a été observé dans le sud de la France en 1975 (Euverte, 1981). Puis son aire de répartition a progressé vers le nord et il est désormais présent sur la très large majorité du territoire national. Son omniprésence est liée à sa capacité de dispersion rapide et à la fréquence de son hôte, le platane commun (Platanus x acerifolia), une des trois espèces les plus plantées en alignement le long des voiries urbaines (Plante & Cité, 2007).
Des nuisances
La connaissance du cycle biologique et de la localisation de l'insecte sur le végétal est essentielle pour positionner les moyens de lutte dans l'espace et le temps.
Deux à trois générations de tigres par an se succèdent selon de la région et les ressources nutritionnelles disponibles (Chauvel, 1988 ; Chapin et al., 2006). À la fin de l'été, les adultes hivernants migrent vers le tronc et s'abritent sous les rhytidomes (= dans l'écorce externe) pour passer l'hiver avant de recoloniser le feuillage du houppier au début du printemps.
Les piqûres nutritionnelles des larves et adultes provoquent des décolorations et la chute prématurée des feuilles. Outre ce préjudice esthétique, le tigre du platane occasionne des nuisances commoditaires (démangeaison, entrée dans les habitations, miellat sur le mobilier urbain et les véhicules, etc.) pouvant entraîner des plaintes de riverains l'été.
Un programme cohérent
Pour rechercher des solutions de lutte biologique contre le tigre du platane, le choix des auxiliaires s'est porté sur des candidats aux potentialités déjà identifiées (Tavella & Arzone, 1986 ; Chaigneau et al., 2002 ; Tarasco & Triggiani, 2006).
Un système automatisé d'analyse d'images couleur a été spécialement conçu pour quantifier les décolorations foliaires. Il permet une analyse précise et fiable des efficacités.
Les résultats obtenus visent à construire une stratégie globale de biocontrôle, faisant appel à l'utilisation combinée de plusieurs auxiliaires.
Trois entomopathogènes
Essais de 2008 à 2010
Suite à des travaux préliminaires réalisés en 2007 (Anomyme, 2007), trois organismes entomopathogènes fournis par la société Koppert, un champignon (Lecanicillium muscarium souche Ve 6, ex-Verticillium lecanii) et deux nématodes (Steinernema spp.) ont été testés :
– à l'automne, sur tronc, après l'entrée en hivernation des tigres sous les rhytidomes ;
– en fin d'hiver, sur tronc, avant la migration ascendante des adultes vers la frondaison ;
– en été, lors du pic de populations donc de nuisances, fin juillet-début août.
Il s'agissait de déterminer l'agent le plus efficace, la période d'application et la dose optimales, ainsi que l'intérêt d'un adjuvant limitant la dessiccation.
De 2008 à 2010, deux types d'essais différents ont été réalisés :
– expérimentations en milieu urbain par des expérimentateurs selon un protocole rigoureux élaboré par les partenaires de PETAAL ;
– essais de valeur pratique, mise en place et observations réalisées par les services techniques des villes concernées par le problème.
Ces essais ont été conduits sur plusieurs communes de France continentale (nord et sud). Les sites ont été choisis en fonction du nombre :
– de tigres sous rhytidomes pour les essais d'hiver [au moins 35 tigres/dm2 de rhytidome, v. seuils de nuisibilité de G. Chauvel (1998)] et sur feuilles pour ceux de l'été ;
– des platanes, il en fallait entre 120 et 200 selon le nombre de modalités.
Les observations ont porté sur le nombre de tigres (vivants et morts)/dm² de rhytidome (Chauvel, 1988) pour les essais d'automne et fin d'hiver, et le nombre de tigres (vivants et morts)/ feuille pour les essais d'été. Pour ces derniers, elles ont été complétées par l'estimation des dégâts foliaires (classes de décoloration, Chapin et al., 2007). De plus, les nématodes étant sensibles à la dessiccation (Galeano, 2009), on a testé l'intérêt d'un adjuvant à base d'huile de colza pour les applications sur tronc.
L'analyse statistique a fait appel à un modèle linéaire généralisé, basé sur une distribution de quasi-poisson (à l'aide des logiciels Statbox 7.1 et R). L'efficacité corrigée est calculée en suivant la formule de Henderson et Tilton (1955).
À partir de 2010, les Steinernema spp. seuls en lice
À l'issue des travaux, il n'y avait aucune différence statistique d'efficacité entre les nématodes et le champignon entomopathogène. En 2010, on a continué avec les Steinernema et abandonné L. muscarium. Ce choix s'appuie sur les exigences bioclimatiques des espèces.
La dose et l'adjuvant
En 2008, les deux doses de nématodes testées en application hivernale sur alignements de platane, 0,6 N et 1 N (N = 2 500 nématodes/ ml) étaient celles employées en application foliaire sur cultures sous abri. Aux deux doses, les niveaux d'efficacité étaient insatisfaisants. Le volume de bouillie étant faible (5 à 10 l/tronc ou 5 à 15 l/houppier, selon la taille de l'arbre), l'hypothèse d'un effet dose a été posée. La dose 3 N a donc été retenue pour les essais suivants.
Pour l'ensemble des essais, les différences ne sont pas significatives du fait de fortes variabilités et du rôle de certains facteurs notamment climatiques. Les meilleures efficacités étant toujours obtenues avec l'association « nématodes + adjuvant », celle-ci a été retenue en 2011.
Période d'application : abandon du traitement automnal
Dans les conditions des quatre essais conduits à l'automne en 2008 et 2009, l'efficacité des nématodes n'a pu être démontrée. Des températures nocturnes et diurnes trop basses aux périodes d'application (octobre) sont la principale hypothèse. Aussi, l'option d'intervenir à cette période n'a pas été retenue.
Fin d'hiver : concilier température douce et migration pas commencée
Lors des 4 essais préliminaires en 2007, les applications de fin d'hiver de nématodes avaient montré une bonne efficacité, la mortalité corrigée allant jusqu'à 81,6 % (Anonyme, 2007).
En 2009 (2 essais) et 2010 (3 essais), la variabilité des résultats a nuancé ce constat. Il est probable que l'action des auxiliaires soit très affectée par les conditions climatiques lors des applications et par la migration des tigres.
En effet, lors des applications réalisées en mars avant la migration, les températures supérieures à 10 °C ont été très rarement atteintes. Et lors des applications début avril (températures optimales), la majeure partie de la population de tigres avait migré, donc a échappé aux nématodes.
Ceci témoigne des difficultés de réunir les facteurs clés d'efficacité d'un traitement sur tronc et charpentières, à savoir la présence des tigres sous rhytidomes et les conditions climatiques nécessaires à l'action des nématodes.
Il a été décidé de continuer à tester les applications de fin d'hiver, avant le début de la migration des tigres mais sous réserve de conditions favorables (température supérieure à 10-12 °C pendant les 3 heures suivant l'application).
Et les applications estivales ?
L'été 2010, deux applications d'été ont été réalisées à deux semaines d'intervalle pour les expérimentations ; dans les essais de valeur pratique, une seule application a été effectuée quand les villes le jugeaient opportun (souvent suite à des plaintes liées aux nuisances).
L'analyse statistique des données issues de ces applications estivales n'a pu être réalisée. Les efficacités corrigées oscillent entre 25 et 96 % selon les sites et le délai entre traitement et observation (Tableaux 1 et 2), sauf à Strasbourg où l'application a été effectuée début septembre. Le début de la migration des tigres vers les rhytidomes à cette période explique probablement l'absence d'efficacité sur ce dernier site.
Bilan provisoire
La mise en œuvre des expérimentations a rencontré des difficultés méthodologiques (nombre de platanes, niveau d'infestation, type d'alignement, quantité de rhytidomes, etc.) liées aux contraintes du milieu urbain.
De plus, les conditions climatiques différentes d'une année à l'autre ont rendu difficile la reproductibilité des résultats. Les conditions climatiques (température et humidité dans les heures suivant l'intervention) et la maîtrise de l'application (dérive, lessivage, qualité de pulvérisation) semblent cruciales.
Un prédateur
Nous avons aussi étudié la capacité prédatrice de l'insecte prédateur indigène Chrysoperla lucasina (Lacroix, 1912) vis-à-vis du tigre du platane. Il s'agissait de déterminer le potentiel et les modalités d'emploi (stade et dose d'apport) contre les populations du ravageur in situ.
Test in vitro sur larves
Des études préliminaires ont montré que C. lucasina consommait préférentiellement les larves de C. ciliata, pour seulement 10 % des adultes (Çaldumbide, non publié), et aucun œuf.
Pour évaluer la capacité prédatrice de C. lucasina, une expérience a été mise en place en laboratoire en conditions semi-contrôlées : température de 20-25 °C, hygrométrie relative de l'ordre de 50 % et photopériode naturelle passant de 13:11 à 12:12 [L:D].
Une larve néonate de C. lucasina, issue d'œufs fournis par la société If Tech, et un nombre non limitant de larves de quatrième et cinquième stades de tigre du platane (prélevées sur des arbres infestés à Angers) sont placées dans une même boîte de Petri. Un relevé quotidien de toute la population de chaque boîte permet d'effectuer un suivi précis de la consommation des chrysopes durant leur développement larvaire. La modalité traitement est répétée dix fois et la modalité témoin répétée une fois.
Résultats : capacité de prédation
Sur les 10 larves de C. lucasina observées, deux sont mortes après quatre jours et les huit autres ont effectué la totalité de leur développement larvaire (14 jours) en consommant uniquement des larves âgées de C. ciliata. Parmi les huit, cinq ont atteint le stade adulte 15 à 16 jours après nymphose, soit un stade nymphal d'une durée moyenne de 15,2 j à 20-25 °C. Dans nos conditions d'essais, et sur les 12 premiers jours (avant la première nymphose), les larves de C. lucasina (n = 8) ont consommé en moyenne entre 4,4 (± 2,1) et 28 (± 3,4) larves de tigre par jour selon le stade larvaire du prédateur (L1 à L3), ce qui fait 150 larves sur l'ensemble du cycle larvaire des chrysopes.
À l'instar d'autres études (Nicoli et al., 1991 ; Malais et Ravensberg, 1992 ; Chaigneau et al., 2002), le stade L3 de C. lucasina est celui ayant la capacité prédatrice la plus importante, jusqu'à 38 larves consommées par jour.
Essai en ville en 2010
Un essai visant à confirmer la prédation in situ et évaluer la dispersion des chrysopes au sein de la frondaison a été réalisé sur l'avenue Jeanne d'Arc à Angers (photo p. 33). Cette voie orientée est-ouest comprend un quadruple alignement de platanes. Taillés d'abord en tête de chat puis conduits en forme semi-libre, ils subissent aujourd'hui une taille mécanique architecturée (rideau) tous les deux ans, la dernière datant de 2009. Les plus grands sujets, d'environ 15 m de haut, ont un fût haut de 5 à 6 m.
Les deux modalités testées (témoin/traitée) sont répétées cinq fois chacune (un arbre = une répétition, soit 10 arbres au total). Le lâcher a été effectué le 1er juin 2010, soit 6 semaines après la migration vers la frondaison, en ciblant les larves de 1re génération.
Un dispositif de lâcher breveté par la société If Tech (tube cartonné contenant 120 à 150 œufs de chrysope) a été placé dans la modalité traitée. Un tube vide d'œufs a été placé dans la modalité témoin. Un comptage des larves et adultes vivants de tigre du platane a été effectué avant le lâcher (J0) et quinze jours après (J+15) à 0, 1 et 2 m du point de lâcher.
La correction des mortalités a été réalisée à l'aide de la formule de Schneider-Orelli (1947). L'analyse statistique (à l'aide du logiciel R) du nombre d'individus par feuille et de la mortalité fait appel à un modèle linéaire généralisé, basé sur une distribution de quasi-poisson. Pour le second type de données, la mortalité moyenne observée dans le témoin a été appliquée aux effectifs initiaux de la modalité traitée, puis comparée à celle observée quinze jours après le traitement dans la modalité traitée.
Prédation et dispersion sur les platanes
L'analyse des effectifs avant traitement montre une influence significative de l'arbre, de la distance et de la modalité (P < 0,05). Le nombre réduit de répétitions n'a pas permis de prendre en compte l'hétérogénéité naturelle importante de l'infestation au sein d'un arbre et entre les arbres. L'infestation significativement plus importante dans la modalité témoin que dans la modalité traitée à J0 nous empêche de conclure formellement sur l'efficacité du lâcher.
Cependant, si l'on prend en compte les mortalités corrigées en fonction de celle observée dans le témoin, on constate 15 jours après le lâcher une réduction significative des populations à 0 et à 1 m (P < 0,05) ainsi qu'à 2 m (P < 0,1) dans la modalité traitée. Les réductions sont respectivement d'environ 70, 55 et 35 % (Tableau 3).
Il semble que le lâcher d'œufs de C. lucasina agisse sur la densité de population de C. ciliata au moins jusqu'à 2 m du point de lâcher.
Bilan provisoire
L'essai in vitro a montré la forte capacité prédatrice des larves de C. lucasina vis-à-vis des larves âgées de tigre du platane. En moyenne, C. lucasina peut consommer près de 150 larves âgées de tigre du platane durant son développement larvaire, soit environ 14 jours à 20-25 °C (Osman & Selman, 1996).
Les conditions de l'essai in situ n'ont pas permis de conclure avec certitude sur l'efficacité des chrysopes, avec cependant une forte réduction des populations de tigre dans la modalité traitée et une dispersion du prédateur jusqu'à 2 m des lâchers. Des essais complémentaires seraient nécessaires pour tenir compte de l'hétérogénéité de l'infestation.
C. lucasina semble donc un prédateur efficace. Mais sa préférence alimentaire pour les larves et sa relative dispersion (comblée par plusieurs points de lâchers) dans le platane suggèrent de l'utiliser dans le cadre d'une stratégie la combinant à d'autres organismes s'attaquant aussi bien aux larves qu'aux adultes et aux modalités d'apport différentes.
Concernant la période d'application, d'après le cycle de vie de C. ciliata (Euverte 1981 ; Zangheri 1986 ; Chauvel, 1988), on considère qu'un lâcher de C. lucasina peut avoir lieu environ six semaines après la migration ascendante des adultes de tigre hivernants vers la frondaison des platanes. Ceci vise à réduire la pression parasitaire en diminuant les populations de larves de première génération (Priéto, 2005).
2011, stratégie combinée
Les travaux portant sur l'efficacité propre des organismes entomopathogènes et des prédateurs ont permis de proposer une stratégie globale de protection biologique par augmentation. L'objectif du travail mené en 2011 est de montrer l'efficacité de combiner plusieurs applications et d'identifier les plus stratégiques.
Essais de valeur pratique dans sept sites de six villes
Il a semblé nécessaire de cibler l'ensemble des phases d'hivernation et d'activité de C. ciliata. Une stratégie combinée a donc été testée dans six villes partenaires (sept sites) dans le cadre d'essais de valeur pratique.
Le choix des sites s'est porté sur des alignements de platanes très infestés les années précédentes. Pour chaque site, deux modalités (témoin/traitée) ont été répétées deux fois.
Une parcelle élémentaire contenant dix arbres (six observés et quatre arbres de garde pour contrôler la dérive et l'effet bordure), chaque essai portait sur quarante arbres.
En amont, un réseau national d'observation a été mis en place afin d'identifier précisément la période de migration de C. ciliata. Ce suivi permettra de valider un modèle de degrés-jour (Herms, 2004) tenant compte aussi de la photopériode, à partir des températures seuils de développement du ravageur (Kim et al., 1999 ; Ju et al., 2011). Le but est de prédire l'activité du tigre et de la corréler au stade phénologique du platane (Miller et al., 2001).
Stratégie à quatre interventions
Les quatre interventions sont (Figure 1) :
• Traitement 1 contre les adultes hivernants à l'aide de Steinernema spp. sur tronc et charpentières avant migration de fin d'hiver. Effectué en mars, températures supérieures à 10-12 °C.
• Traitement 2 contre les adultes hivernants à l'aide de Steinernema spp. sur feuillage après la migration ascendante vers la frondaison. Effectué courant mai.
• Traitement 3 contre les larves de première génération sur le feuillage à l'aide d'œufs de C. lucasina. Effectué six semaines après migration du tigre soit en juin.
• Traitement 4 à l'aide de Steinernema spp. fin juillet-début août contre les individus présents sur feuillage.
À la fin de la campagne, des feuilles de platane ont été prélevées pour analyse à l'aide du système automatisé d'analyse d'image couleur mis au point dans le cadre du programme (Clément et al., 2011). L'analyse était encore en cours lors de l'écriture de ces lignes fin octobre.
Résultats sur les populations
Les résultats sur populations de tigre sont :
– une efficacité significative des nématodes en fin d'hiver sur tronc et charpentières (réduction de 60 % en moyenne des populations de tigre 7 jours après application des nématodes) ;
– une efficacité significative de la combinaison des trois interventions de mai à début août (mesurée 7 jours après le dernier traitement) ; le traitement de mai a permis une réduction moyenne des populations de 45 %, l'apport de chrysopes n'a pas eu d'effet significatif sauf à Antony (50 %) et l'apport de nématodes en été a eu un effet significatif sur des populations faibles (les conditions climatiques de 2011 n'ont pas permis d'observer de pic de population estival comme il s'en manifeste habituellement).
Conclusion
Les travaux conduits depuis 2008 ont permis de dégager les composantes d'une stratégie combinée pour gérer les populations de tigre du platane et contrôler ses nuisances. Deux espèces de nématodes entomopathogènes (Steinernema spp.) et l'insecte prédateur indigène C. lucasina ont été retenus.
De bonnes pratiques, c'est-à-dire la maîtrise des conditions d'intervention et le positionnement judicieux de chacune d'entre elles, sont essentielles pour garantir l'action de ces agents de biocontrôle.
À l'issue du programme PETAAL, il sera possible de proposer un itinéraire technique compatible avec le cadre réglementaire de la protection phytosanitaire en espaces verts et correspondant aux attentes techniques et économiques des professionnels.
<p>* Agrocampus Ouest - Centre d'Angers INHP 2, rue Le Nôtre, 49045 Angers cedex 01. thibaut.verfaille@agrocampus-ouest.fr, bruno.jaloux@agrocampus-ouest.fr</p> <p>** Koppert France - 14, rue de la Communauté Parc d'Activités de Viais, 44860 Pont-Saint-Martin. mpiron@koppert.fr</p> <p>*** Plante & Cité - 3, rue Alexander-Fleming 49006 Angers cedex 01. caroline.gutleben@plante-et-cite.fr</p> <p>**** If Tech - Centre Horticole Floriloire 3, rue des Magnolias, 49130 Les Ponts-de-Cé. contact@iftech.fr</p> <p>*** ** Fredon Provence-Alpes-Côte-d'Azur, Antenne du Var 224, rue des Découvertes, 83390 Cuers. annemaury.fredon@orange.fr</p> <p>*** *** Université d'Angers - Laboratoire LISA, Institut Universitaire de Technologie, 4, bd Lavoisier, BP 42018, 49000 Angers cedex. alain.clement@univ-angers.fr</p>
Tableau 1 - Efficacité corrigée [après les 2 traitements (T1 et T2) avec les nématodes] lors des essais expérimentaux d'applications d'été en 2010.
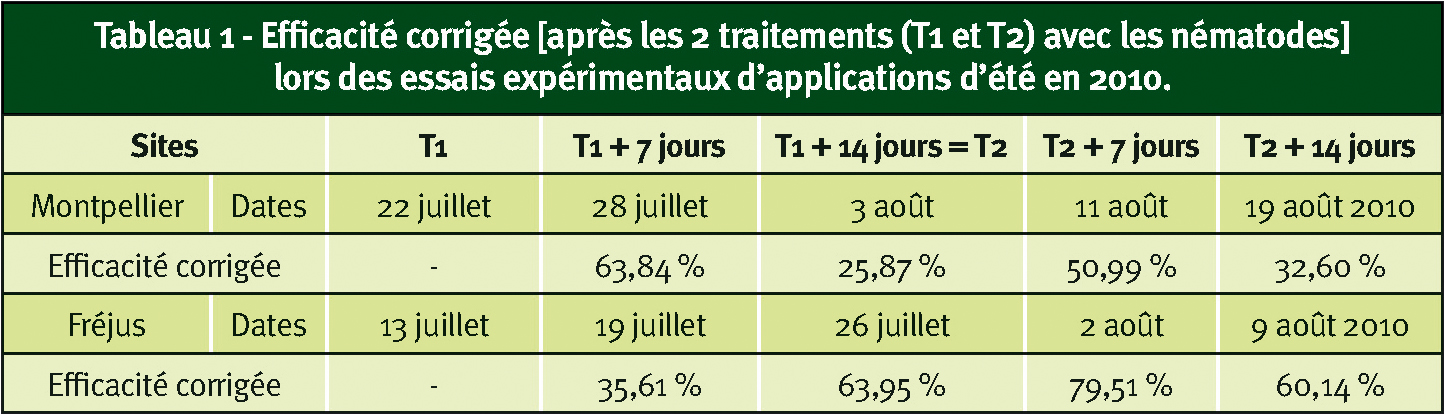
Tableau 2 - Efficacité corrigée (7 et 19 jours après le traitement (T) avec les nématodes) et appréciation de l'efficacité par les villes lors des essais en été 2010.
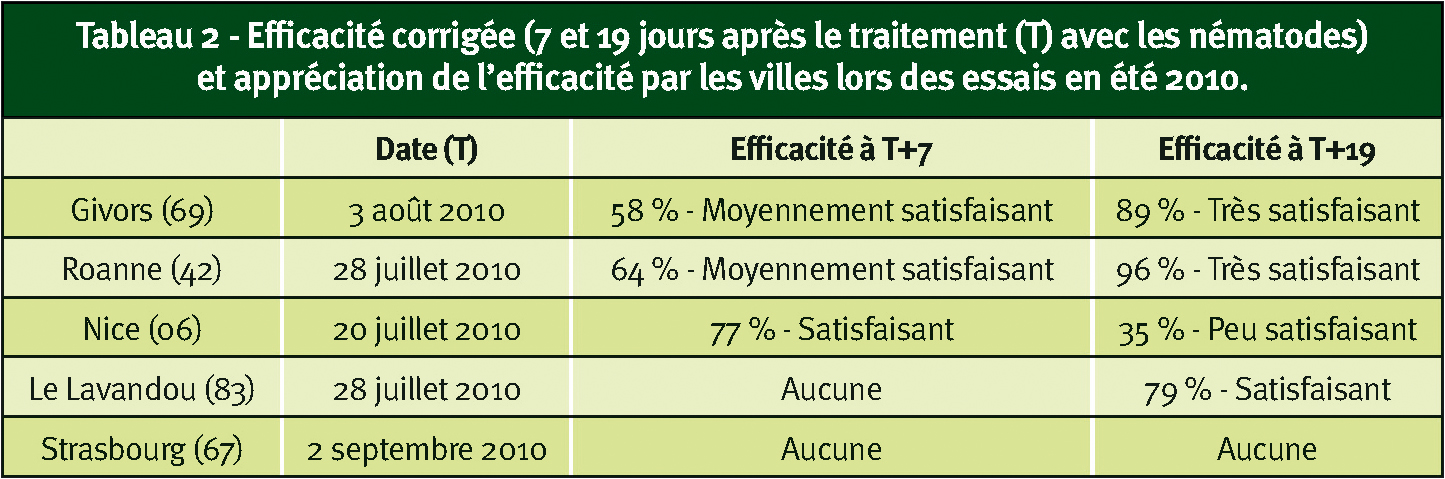
Tableau 3 - Mortalité des individus (larves et adultes) de C. ciliata 15 jours après un lâcher d'œufs de C. lucasina à différentes distances du point d'apport.
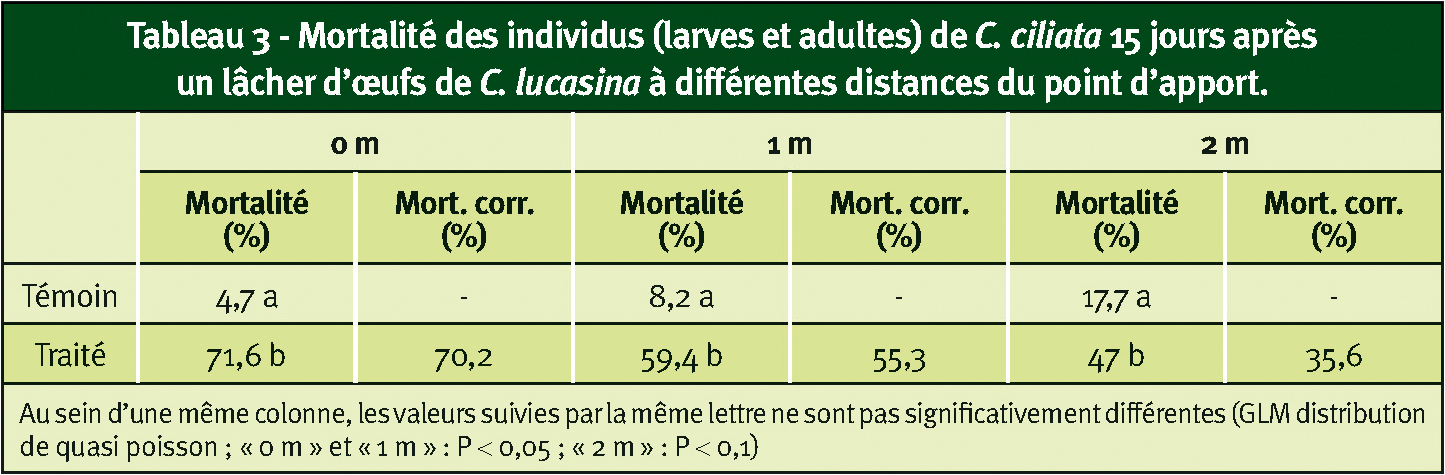
Figure 1 - Stratégie globale de contrôle biologique des populations de tigre du platane testée en 2011 (représentation schématique du cycle de vie de C. ciliata).


ph. T. Verfaille

