
 Imprimer
Imprimer par SPME du maïs dans l'espace de tête d'un maïs cultivé au champ. Les COV seront ensuite identifiés au laboratoire. Photo : E. Leppik, Inra")
Prélèvement de COV (composés organiques volatils, ou « odeurs ») par SPME du maïs dans l'espace de tête d'un maïs cultivé au champ. Les COV seront ensuite identifiés au laboratoire. Photo : E. Leppik, Inra

Pyrales du maïs utilisés lors des tests : à gauche, la femelle, à droite, le mâle. Photo : E. Leppik, Inra
 du maïs. Photo : M. Decoin")
En plein jour comme ci-dessus, les pyrales adultes sont rarement présentes dans le champ de maïs. Les femelles s'y rendront le soir pour pondre, guidées par les COV (les « odeurs ») du maïs. Photo : M. Decoin
La pyrale du maïs Ostrinia nubilalis (Hübner) (Lepidoptera, Crambidae) est le principal ravageur du maïs dans l'hémisphère nord.
Les études du comportement ont montré que ce ravageur utilise des signaux chimiques pour localiser et discriminer sa plante-hôte (Leppik et al., 2012). Est-il possible d'utiliser ces signaux pour leurrer la pyrale ?
À la découverte des COV
Signatures chimiques
Le maïs (Zea mays), comme toutes les plantes vasculaires, émet des composés organiques volatils (COV) qui sont des métabolites secondaires (Dudareva et al., 2000). Chaque biotope et chaque parcelle cultivée émettent un mélange de COV spécifiques constituant un paysage chimique original (Leppik et al., 2014) et propre à ce biotope.
Les insectes herbivores qui ont coévolué avec les plantes (Schoonhoven et al., 2005) utilisent des signatures chimiques constituées d'un assemblage spécifique de COV pour reconnaître leur plante-hôte (Beyaert et al., 2014).
En particulier, les pyrales adultes ne restent pas dans les champs de maïs durant la journée. Elles se réfugient dans des zones herbacées et humides où elles s'accouplent (Showers et al., 1976). Au crépuscule, seules les femelles fécondées vont se diriger vers les champs de maïs pour y pondre.
Une perspective pour la protection ?
À côté des phéromones sexuelles qui sont utilisées pour la surveillance des vols de ravageurs, l'utilisation des COV pour contrôler les populations des ravageurs est un domaine qui peut présenter une forte potentialité.
Une récente étude a démontré que les pyrales fécondées sont attirées en tunnel de vol par un mélange de nonanal et décanal (Molnár et al., 2015), mais il reste encore à démontrer au champ l'efficacité de ce mélange.
Nous présentons ici une étude au cours de laquelle nous avons :
- identifié les odeurs impliquées dans la reconnaissance de la plante-hôte ;
- étudié la détection des composés identifiés par électro-antennographie (EAG) ;
- déterminé un mélange attractif en tunnel de vol pour les femelles fécondées.
Nos résultats sont présentés et discutés ici dans la perspective d'une application pour maîtriser les populations de la pyrale.
Du champ à l'antenne
Prélèvement des odeurs au champ
Les COV sont prélevés dans l'espace de tête (50 × 30 cm) d'un maïs cultivé au champ, au stade 4 feuilles (Grignon, Yvelines, France) (48 °85' N, 1,9 °68' E) sur la variété Troubadour. Les prélèvements sont effectués pendant 2 heures le jour de 13 h 00 à 15 h 00, puis durant deux heures également en début de nuit, soit de 22 h 00 à 00 h 00 (dispositif visible sur la photo ci-dessus).
Analyse des COV
Les fibres SPME (solide phase micro extraction) utilisées sont des divinylbenzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS- 50/30 µm. Supelco).
Les fibres sont désorbées dans l'injecteur d'un chromatographe en phase gazeuses (Scion 436) couplé au détecteur masse simple quad SQ TM (Bruker).
Les COV sont séparés sur une colonne Rxi-5ms (Restek, France) 30 m × 0,32 mm I. D., épaisseur de film 0,5 µm. Le four de la colonne est programmé de 50 °C à 300 °C par pas de 8 °C/minute avec l'hélium comme gaz vecteur. Les spectres de masse sont obtenus en mode impact électronique (70 eV).
Les COV sont identifiés grâce à leurs spectres de masse et leurs indices de rétention (RI). Les RI sont calculés en utilisant les n-alcanes C10 à C24, élués dans les mêmes conditions que les échantillons. Les spectres des composés élués et les RI sont comparés à ceux de la bibliothèque du laboratoire et de la bibliothèque NIST 2011 par le biais de AMDIS32.
Étude électrophysiologique de la perception des odeurs par la pyrale
Les enregistrements EAG (électro-antennographie) ont été effectués à température ambiante sur les pyrales âgées de 3 à 4 jours selon la méthode décrite par Renou et al., 1986 (Figure 1). L'analyse des réponses EAG est effectuée à l'aide du logiciel pClamp 10 (Molecular Devices).
Chaque composé a été testé après un repos de 1 minute, deux fois sur l'antenne et les réponses moyennes sont utilisées pour les résultats.
Formulations des attractifs
Quatre formulations différentes contenant trois et six molécules candidates ont été testées en tunnel du vol et confrontées à un témoin : une plante de maïs au stade 4 à 6 feuilles.
Tests de comportement de la pyrale en tunnel vis-à-vis des attractifs
Les tests sont réalisés sur des insectes dont les larves ont été prélevées sur la plante et qui ont terminé leur développement sur du milieu semi-artificiel. Les femelles fécondées pondent au début de la scotophase (période d'obscurité du cycle jour/nuit), et tous les tests de laboratoire ont respecté ce rythme d'activité. Le tunnel du vol est parcouru par un flux d'air contrôlé et laminaire. Une très faible lumière rouge permet l'observation des comportements qui sont enregistrés par une caméra infrarouge placée au-dessus du tunnel (Figure 2).
Pour chaque individu testé, les paramètres de l'attraction notés sont : l'activation, le vol orienté dans le flux d'air odorisé et le vol autour de la source.
Résultats
Les COV émis par le champ de maïs
Le bouquet odorant émis par le maïs, composé d'une dizaine de COV, est relativement pauvre. C'est un mélange à base d'odeurs vertes, de monoterpènes et de sesquiterpènes. Il est à noter que les ratios changent entre le jour et la nuit (Figure 3).
Ainsi, les sesquiterpènes (α-copaène, β-caryophyllène, δ-cadinène) sont majoritairement émis pendant le jour, tandis que les monoterpènes (α-pinène, p-cymène, limonène, linalool) et certaines odeurs vertes comme cis-3-hexenyl acétate et cis-3-hexenol sont majoritairement émis pendant la nuit.
Résultats du test d'électro-antennographie
De façon générale, les femelles détectent mieux les COV du maïs que les mâles. Dans le détail, les deux sexes détectent très bien certains monoterpènes (COV 4, COV 6, COV 9) et sesquiterpènes (COV 13), mais pour les COV 1 (une odeur verte) et COV 5 (un monoterpène), les femelles sont deux fois plus sensibles que les mâles (Figure 4).
Comportement de la pyrale vis-à-vis de l'odeur synthétique du maïs
Une série de tests non présentés ici a permis de sélectionner un mélange pertinent qui induit les mêmes comportements d'attraction que ceux suscités par le maïs (voir Figure 5).
Un profil odorant spécifique
La femelle de la pyrale, qui pond au début de la nuit, utilise les COV émis par le maïs pendant cette période pour localiser la parcelle où elle va pondre.
Les résultats montrent clairement que les signaux chimiques produits par le maïs sont différents entre le jour et la nuit (Leppik et Frérot, 2014). L'ensemble de ce profil odorant est relativement pauvre en COV, mais la composition tant qualitative que quantitative est spécifique. Le profil odorant du maïs au champ est différent du profil des plantes cultivées en pot en conditions du laboratoire.
L'émission des COV par les plantes est dépendante des conditions biotiques : du métabolisme, du stade de développement, de la présence des bioagresseurs, etc., et de facteurs abiotiques comme le climat, le cycle circadien, le sol, le déficit hydrique, la pollution, etc.) (Holopainen et al., 2010 ; Jakobsen et al., 1994 ; Loreto et al., 2010).
Ce point peut expliquer que nos résultats sur la composition de l'attractif diffèrent de ceux décrits par Molnár et al., 2015, bien que nous confirmions la présence des deux constituants décrits comme attractifs : nonanal et décanal. Ces produits seront évalués sur le terrain, soit seuls, soit en complément de notre attractif.
La reconstitution de l'odeur naturelle pertinente avec les composés de synthèse sélectionnés sur la base de la réponse olfactive permet d'obtenir dans les conditions du laboratoire des comportements identiques à ceux obtenus avec la plante (Figure 5). Ces résultats confirment les observations antérieures montrant que la localisation de la plante-hôte était assurée par reconnaissance olfactive.
Les tests d'attractivité sur le terrain permettront de savoir si les autres ravageurs du maïs peuvent être attirés par ce mélange. L'attractif de synthèse est un nouvel exemple potentiel de l'intérêt des kairomones dans la protection des plantes, pour la surveillance du ravageur et la lutte directe contre les femelles fécondées.
Dans ce dernier cas, on pourrait supposer la création de zones où les pontes seraient concentrées, ou encore sélectionner des variétés du maïs qui n'émettent pas ces odeurs attractives. L'utilisation des COV pourrait être à l'avenir un moyen de lutte contre les ravageurs et une alternative supplémentaire aux insecticides.
Fig. 1 : Premier test, réaction des antennes

Photo : E. Leppik, Inra
Poste d'électro-antennographie pour enregistrer et étudier la détection des COV par les antennes de la pyrale : c'est un indice de la perception d'une odeur.
Fig. 2 : Deuxième test, comportement d'attraction
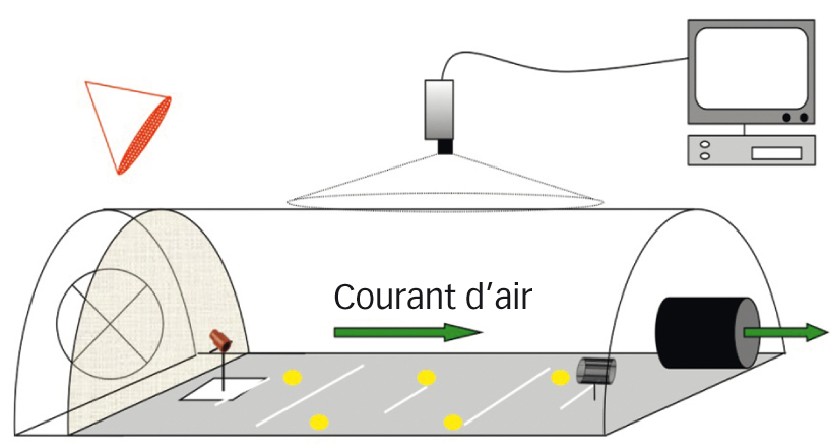
Tunnel de vol utilisé pour étudier le comportement de pyrales vis-à-vis des odeurs du maïs, naturelles ou de synthèse.
Fig. 3 : Profils chromatographiques de l'espace de tête prélevé par SPME in situ sur des maïs pendant le jour et la nuit
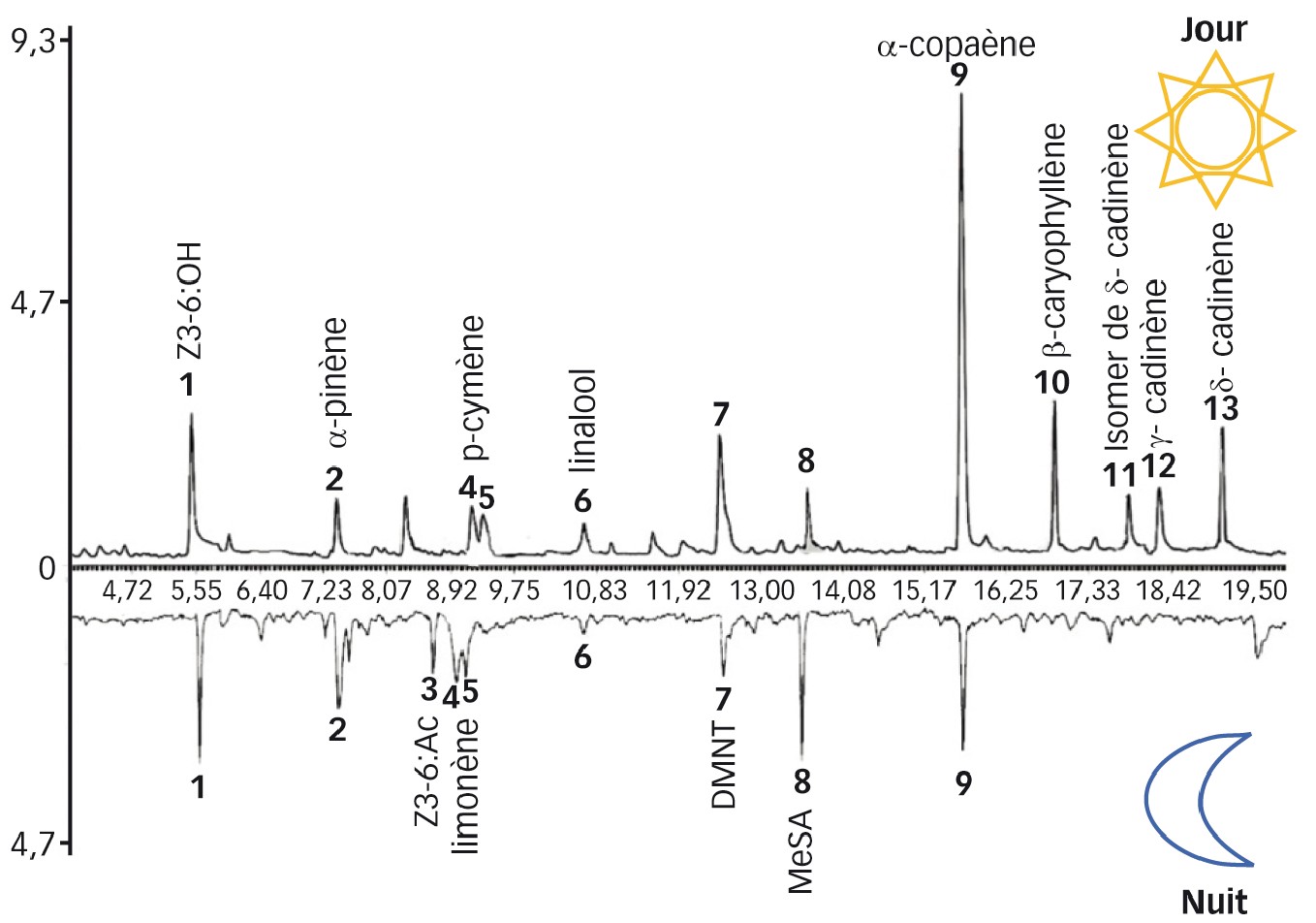
Il est visible que les proportions des COV émises diffèrent entre les deux périodes.
Fig. 4 : Les réponses EAG des antennes de pyrale stimulées par les COV identifiés par SPME-GC-MS
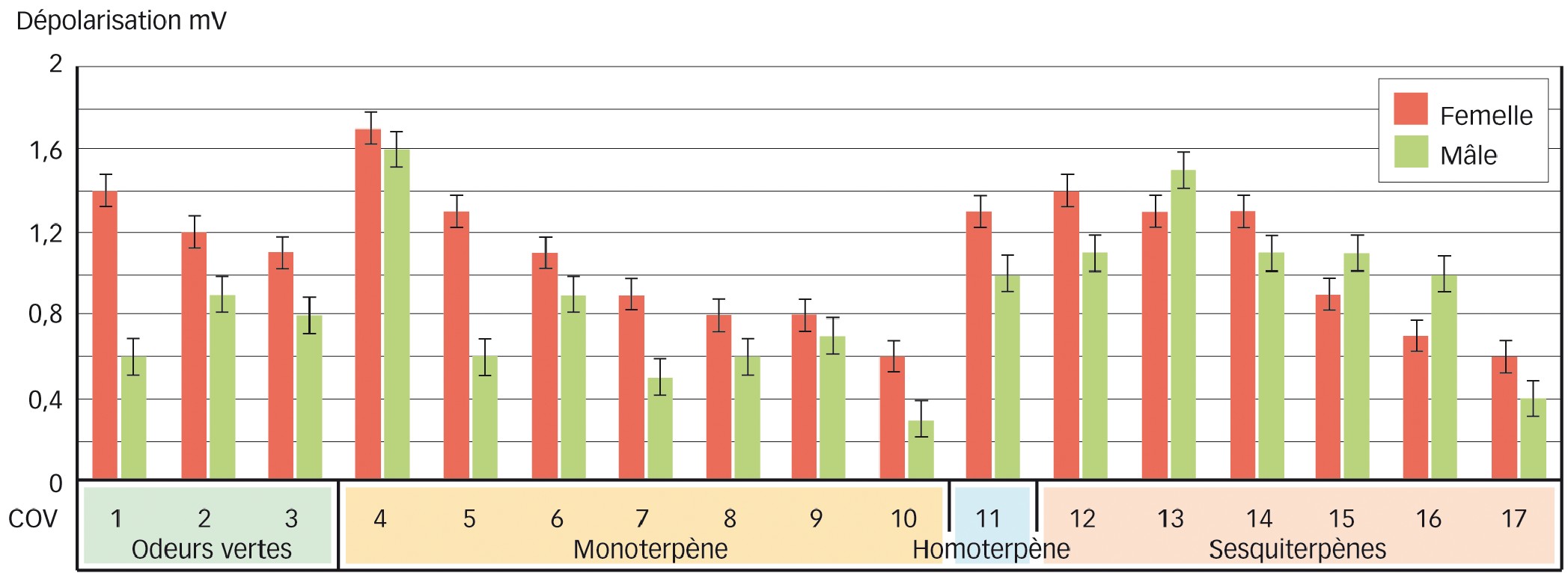
Les réponses, indice de perception de chacun des COV (chacune des odeurs), diffèrent entre mâles et femelles.
Fig. 5 : Vrai maïs ou attractif ?

Pourcentage d'attraction des femelles fécondées de pyrales stimulées par le maïs et par quatre attractifs de synthèse. Un de ces derniers égale significativement le vrai maïs.
REMERCIEMENTS
REMERCIEMENTS à Philippe Lucas pour la mise à disposition du poste d'électro-antennographie, et à Arvalis et la FNPSMS qui ont financé la recherche.

