 Imprimer
Imprimer
Photo : Pixabay

Photo : Pixabay

Femelle Drosophila suzukii sur un substrat de fraise. Cette mouche apprécie de nombreux fruits rouges, tels que la cerise et la fraise, et pond dans les fruits mûrs et encore fermes. Photo : S. Lebreton

Composition des quatre substrats de ponte testés
La drosophile à ailes tachetées, Drosophila suzukii Matsumura, est une espèce invasive qui s'est implantée en Europe et en Amérique du Nord en provenance d'Asie en 2008, et s'est depuis répandue sur l'ensemble de ces continents (Fraimout et al., 2017).
Comment D. suzukii attaque les fruits mûrs mais intacts
Un atout morphologique
Contrairement aux autres espèces de drosophiles qui pondent sur des fruits abîmés, D. suzukii s'attaque préférentiellement aux fruits mûrs mais encore fermes.
Cette caractéristique comportementale est en partie due à une modification de la structure de son ovipositeur (appendice à l'aide duquel les femelles pondent leurs oeufs), lui permettant de percer la peau des fruits intacts (Atallah et al., 2014).
De plus, D. suzukii est une espèce généraliste qui s'attaque à une large gamme de petits fruits, tels que la fraise, la framboise, la cerise, la myrtille ou bien encore le raisin (Walsh et al., 2011).
Toutes ces caractéristiques font de cette drosophile l'un des ravageurs principaux sur les cultures de fruits de nombreux pays (Bolda et al., 2010 ; Lee et al., 2011).
Des caractéristiques sensorielles
Si l'adaptation morphologique de son ovipositeur explique pourquoi D. suzukii est capable de pondre sur des fruits intacts, la préférence pour ces derniers s'explique par une modification de sa perception sensorielle (Karageorgi et al., 2017).
Par exemple, D. suzukii est beaucoup moins sensible à la dureté du substrat sur lequel elle pond, en comparaison avec des espèces proches, comme Drosophila biarmipes Malloch ou bien Drosophila melanogaster Meigen. Ces dernières espèces, connues pour s'attaquer aux fruits abîmés, choisissent toujours les substrats les plus mous (Karageorgi et al., 2017).
Autre modalité sensorielle affectée : l'olfaction. Nous avons ainsi pu mettre en évidence récemment que l'odeur de fraises mûres suffit à elle seule pour stimuler la ponte de D. suzukii, ce qui n'est pas le cas de D. melanogaster (Karageorgi et al., 2017).
Résultats antérieurs
Des lignées à récepteurs olfactifs « manipulés »
Les drosophiles, tout comme les autres insectes, détectent les composés chimiques volatils par le moyen de deux types de récepteurs : les récepteurs olfactifs de type OR (olfactory receptors) et les récepteurs ionotropiques (IR = ionotropic receptors) (Benton et al., 2009 ; Larsson et al., 2004).
Pour aller plus loin dans l'étude de la perception olfactive chez D. suzukii, nous avons généré des lignées mutantes et transgéniques de cette espèce présentant des systèmes olfactifs manipulés.
Deux types de signaux
Ceci nous a permis de montrer que les récepteurs olfactifs de type OR sont responsables de la perception des composés volatils émis par les fraises mûres qui stimulent la ponte de cette mouche (Karageorgi et al., 2017).
Cependant, nous avons noté un comportement inattendu de la part des mouches anosmiques (ne percevant pas d'odeur) pour les composés détectés par les récepteurs de type OR. En effet, ces mouches se comportent normalement lorsqu'elles sont mises en contact direct avec les fruits mûrs (Karageorgi et al., 2017).
Ce résultat suggère que des signaux perçus par contact, donc probablement des signaux gustatifs, sont eux aussi capables de stimuler la ponte chez D. suzukii et peuvent compenser la perte partielle de l'odorat.
Nous nous sommes par conséquent intéressés dans cette étude à la part que prend chacun de ces deux types de signaux (olfactif et gustatif) dans la stimulation de la ponte, ainsi que la manière dont ils interagissent pour moduler le comportement de D. suzukii et D. melanogaster.
Matériel et méthode
Élevage de deux lignées sauvages, chacune d'une espèce différente
Deux lignées sauvages - « Oregon R » (D. melanogaster) et « Alpes-Maritimes, AM » (D. suzukii) - ont été utilisées pour réaliser cette étude. Les deux stocks étaient maintenus dans des tubes standard communément utilisés pour l'élevage des drosophiles contenant de la nourriture NutriFly(1). Dans le cas de D. suzukii, une bande de papier Whatmann était ajoutée dans chaque tube afin de permettre la pupaison.
Dans les deux jours suivant l'émergence, les mouches ont été transférées dans un nouveau tube pendant sept à dix jours avant le début de l'expérience.
Dispositif expérimental : les « boîtes gigognes »
Environ 6 h avant le début de l'expérience, les mouches étaient anesthésiées à l'aide de CO2, puis les femelles de l'une ou l'autre des deux espèces étaient placées dans un nouveau tube, par groupe de dix. Chaque groupe de femelles était ensuite transféré dans une boîte en Plexiglas (12 × 6 × 4 cm) dans laquelle se trouvaient deux boîtes de Petri concentriques où les mouches avaient la possibilité de pondre (Figure 1).
Le dispositif de ponte était constitué d'une petite boîte de Petri (Ø 35 mm) placée au centre d'une boîte plus grande (Ø 55 mm). La petite boîte située au centre était recouverte d'une grille métallique permettant aux mouches de sentir les odeurs émanant de cette boîte tout en les empêchant d'avoir un contact direct avec le contenu. Les mouches pouvaient en revanche pondre autour de la petite boîte, sur le substrat présent dans la plus grande (Figure 1).
Les mouches étaient confrontées à l'un des quatre substrats suivants (n = 23 par condition et par espèce) :
- odeurs de fraises fraîches seules (30 %) ;
- saccharose seul (6 %) ;
- odeurs de fraises fraîches (30 %) + sucres (6 %) ;
- fraises fraîches (30 %).
La composition de ces substrats de ponte est décrite dans le Tableau 1 et illustrée dans la Figure 1. De la purée de fraise Sicoly a été utilisée dans cette expérience(2). La purée contenant 20 % de sucre, une dilution de 30 % de cette purée correspond à une concentration de sucre de 6 % dans le substrat de ponte final (le taux de sucres présents dans une fraise est de l'ordre de 5-6 %). Au bout de 24 h, les boîtes de Petri étaient prélevées et le nombre d'oeufs compté.
Analyse statistique
Pour chaque espèce, le nombre d'oeufs pondus a été comparé entre les différents traitements grâce à un modèle linéaire généralisé (GLM) avec l'hypothèse d'une structure d'erreur binomiale négative, suivi d'un test de comparaisons multiples avec une méthode de correction post hoc de type FDR (false discovery rate).
Résultats
Des comportements de ponte différents selon l'espèce
Drosophila melanogaster ne pond presque pas en présence d'odeurs de fraises uniquement (0,48 ± 0,73 oeufs pondus ; Figure 2A). Elle pond significativement davantage sur le substrat sucré (9,65 ± 12,06). Le taux de ponte augmente encore si les odeurs de fraises sont combinées avec le sucre (22,87 ± 22,16) en comparaison avec le sucre seul. Ce taux de ponte est toutefois inférieur au taux de ponte sur la purée de fraises (70,39 ± 29,81).
En ce qui concerne D. suzukii, sa ponte est stimulée à la fois par les odeurs de fraises (28,39 ± 37,13 oeufs pondus) et par le sucre (38,48 ± 40,25 ; Figure 2B). Lorsque les deux stimuli sont combinés, on observe un taux de ponte (67,82 ± 55,76) intermédiaire entre chacun des stimuli seuls et la purée de fraises (108,00 ± 51,57), tout en n'étant pas significativement différent de cette dernière.
Comportement en milieu naturel
Nous savons que, sur le terrain, les comportements de ponte de D. melanogaster et D. suzukii sont radicalement différents. Alors que D. melanogaster pond quasi exclusivement sur des fruits abîmés ou en cours de décomposition, D. suzukii préfère nettement pondre sur des fruits frais ou en cours de maturation.
Dans leur environnement naturel, ces deux espèces sont donc confrontées à des substrats de ponte dont la composition chimique (volatile et non volatile) diffère grandement.
Lors de la maturation, les fruits accumulent du sucre (essentiellement du fructose, ainsi que du glucose et du saccharose dans une moindre mesure) (Woodward, 1972).
Ensuite, lorsqu'il est abîmé, le fruit va être colonisé par des micro-organismes, notamment des levures. Ces derniers, au cours d'un processus de fermentation, vont dégrader les sucres (Berthels et al., 2004) et produire de l'éthanol, puis de l'acide acétique ainsi que divers autres composés chimiques volatils (Mansourian and Stensmyr, 2015).
Par conséquent, il n'est pas surprenant que les deux espèces aient subi, au cours de l'évolution, des modifications de leurs systèmes sensoriels ayant conduit à des divergences comportementales en réponse à certains signaux chimiques (Karageorgi et al., 2017 ; Keesey et al., 2015 ; Pham and Ray, 2015 ; Ramasamy et al., 2016 ; Revadi et al., 2015).
La « mouche du vinaigre », le sucre et la fraise fraîche
Nos résultats confirment que le comportement de ponte de D. melanogaster, dite « mouche du vinaigre », n'est pas stimulé par la présence seule d'odeurs de fraises fraîches. En revanche, ces mêmes odeurs la stimulent si elle a la possibilité de pondre sur un substrat sucré. L'effet de ces odeurs sur la ponte de ces mouches dépend donc du contexte dans lequel elles les détectent.
La purée de fraises fraîches utilisée dans cette étude pourrait s'apparenter à des fruits tout juste abîmés, avant qu'ils ne soient colonisés par les levures. Or les larves se nourrissent de ces micro-organismes qui ont besoin de sucres pour se développer. Ainsi, un substrat sucré pourrait indiquer un terrain propice au développement des levures et par conséquent des larves de drosophiles.
Ceci expliquerait pourquoi D. melanogaster ne répond aux odeurs de fruits qu'en présence de sucre, d'autant plus que ces mouches ont la capacité d'ensemencer elles-mêmes le milieu de levures au moment de la ponte (Becher et al., 2012).
La « suzukii » pond dans tous les cas
D. suzukii, quant à elle, est stimulée par les odeurs de fraises mûres « seules », comme précédemment établi (Karageorgi et al., 2017), mais également par le sucre seul, et enfin par l'association des deux composants. Ceci est en adéquation avec ses préférences de ponte en milieu naturel, à savoir les fruits mûrs ou en cours de maturation.
Synergie... ou pas
Nos résultats suggèrent également que, chez D. melanogaster, le sucre et les odeurs de fruits ont un effet synergique sur la ponte. En effet, le nombre d'oeufs pondus en présence simultanée des deux stimuli (22,87) est deux fois supérieur à la somme des oeufs pondus en présence de chacun des deux stimuli seuls (0,48 + 9,65 = 10,13).
Ce n'est pas le cas de D. suzukii chez qui on observe plutôt un effet additif : l'effet conjoint des deux stimuli (67,82 oeufs pondus en moyenne) est égal à la somme des effets de chacun des stimuli (28,39 + 38,48 = 66,87 oeufs). Les mécanismes moléculaires et cellulaires à l'origine de l'intégration des signaux gustatifs et olfactifs au niveau du système nerveux central pourraient donc être différents chez ces deux espèces.
Autres différences entre les deux espèces
En définitive, chez Drosophila melanogaster, le nombre d'oeufs pondus sur la purée de fraises est bien supérieur au nombre d'oeufs pondus en réponse à la présence simultanée d'odeurs de fraises et de sucre (qui ne représente que 32 % de la réponse sur la purée de fraises). Des signaux gustatifs autres que le saccharose jouent donc un rôle majeur sur la stimulation de la ponte chez cette espèce.
Chez Drosophila suzukii, en revanche, le nombre d'oeufs pondus sur la purée de fraises n'était pas significativement plus élevé qu'en réponse à la présence conjointe des deux stimuli (la ponte sur [odeurs + sucre] représente 63 % de la ponte sur [fraises]). Contrairement à Drosophila melanogaster, il semblerait donc que D. suzukii se base largement sur la présence d'odeurs et la quantité de sucres pour pondre sur un fruit mûr.
Conclusion
Contrairement aux autres espèces de drosophiles comme D. melanogaster, D. suzukii s'attaque préférentiellement aux fruits peu mûrs, ce qui en fait un ravageur majeur des cultures de fruits (Bolda et al., 2010 ; Lee et al., 2011). Cela suggère que les signaux chimiques (olfactifs et gustatifs) utilisés lors de la ponte, l'importance relative de ces signaux ainsi que l'intégration de ces signaux au niveau du système nerveux a changé chez D. suzukii.
Il s'agira maintenant de déterminer quels sont les signaux spécifiques impliqués dans la stimulation de la ponte chez cette espèce, ainsi que les récepteurs olfactifs et gustatifs responsables de leur perception afin de mettre au point des outils efficaces pour lutter contre cette espèce.
(1) http://flystocks.bio.indiana.edu/Fly_Work/media-recipes/germanfood.htm (2) www.sicoly.fr/fruit/produits-fruits-surgeles/purees/puree-fruits/fraise-puree.html
Fig. 1 : Dispositif expérimental

La petite boîte de Petri, au centre du dispositif, est couverte d'une grille : les drosophiles peuvent sentir l'odeur de son contenu mais pas le toucher, ni le goûter, ni y pondre. En revanche, elles ont accès au « boulevard circulaire » que représente la portion du fond de la grande boîte entourant la petite.
Fig. 2 : Nombre d'oeufs pondus par D. melanogaster (A) et D. suzukii (B) sur les différents substrats de ponte
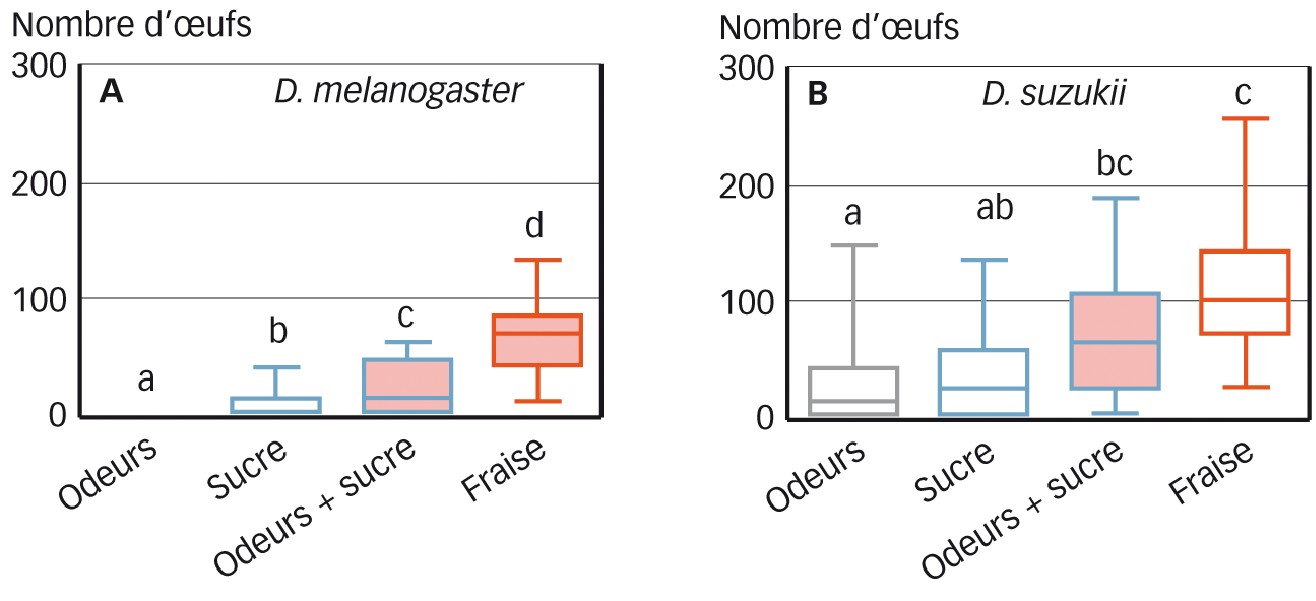
Les lettres au-dessus des histogrammes montrent des différences statistiques entre les différentes conditions au sein de chaque espèce.
REMERCIEMENTS
Ce travail a bénéficié d'une aide du gouvernement français au titre du Programme investissements d'avenir, initiative d'excellence d'Aix-Marseille Université - A*Midex.

